
OBRA Summa totius operis Aristotelis
Libro XII: Biología y Zoología
Titulo IV: El alma sensitiva.
Capitulo III. La audición y los sonidos que producen los animales.
III.1 Las facultades y funciones del alma. La facultad de percepción. La percepción auditiva.
Vuelve a utilizar Aristóteles, respecto al sonido, sus categorías de potencia (ἐνεργείᾳ) y acto (δυνάμει)[1]. Hay unos cuerpos que son silenciosos, en potencia y en acto, y cita entre ellos las esponjas y la lana, y otros que son sonoros en potencia y pueden serlo, o no, en acto. En este caso, está el bronce y cuantos son duros y lisos; pues son capaces de producir un sonido en acto que recorre el medio entre el objeto sonoro y el oído, como consecuencia de un choque. Pues es imposible –continúa escribiendo- que un objeto único produzca el sonido: un objeto ha de golpear y otro ser golpeado. Y en los huecos (κοῖλα) se producen muchas repercusiones (ἀνακλάσει πολλὰς ποιεῖ πληγας μετὰ τὴν πρὠτην) a partir del primer golpe, si los que se producen no pueden salir. [2]
Después de habernos dicho que el medio de transmisión del sonido era el aire, nos dice que también lo es el agua; aunque menos perfectamente (αλλ ἧττον)[3]. Pero que el sonido se produce cuando dos cuerpos duros chocan el uno contra el otro y contra el aire[4].
Luego, nos dice que es el movimiento del aire, como un conjunto único (συνεχἠς καὶ εἰς) el que produce la audición[5]; y que por la inconsistencia del aire, el cuerpo que resuena debe ser liso y extenso, precisamente porque el plano extenso por ambas causas mueve mejor, como una sola masa un conjunto sutil como el aire.
Oímos –prosigue- por el aire[6]. Cuando el aire se mueve, el aire interno encerrado en el oído se mueve a su vez, percibiendo con exactitud todas las variaciones en el movimiento del aire.
En cuanto a la voz –que es el sonido emitido por un animal- solo la poseen los animales que tienen aire dentro de sí para mantener el calor interno necesario para la vida. El órgano de la respiración para Aristóteles es la laringe y su función la de servir, a su vez, al pulmón; pues “es gracias a este órgano que los animales terrestres poseen más calor que los otros” [7].
Con todo, no todo sonido emitido por un animal es voz, sino que es necesario que el choque que produzca la voz corresponda a un movimiento animado y que tenga algún significado:
“Pues la voz, sin duda, es un sonido cargado de significado y no un ruido producido por el aire inspirado” [8].
En la voz se golpea el aire inspirado, contra la pared de la traquea (ὰρτηρίᾳ). No se puede hablar inspirando o expirando: es necesario retener el aire, para lanzarlo contra la traquea[9].
Respecto al oído, las observaciones de Aristóteles son muy sencillas:
“El emplazamiento del órgano auditivo es muy distinto, pues algunos tienen orejas y otros conductos auditivos aparentes”[10]
Afirma que los peces oyen, a pesar de que no tienen ningún órgano aparente de audición[11], porque huyen de los ruidos violentos, como los que producen los remos de las trirrenes; “pues un ruido por débil que sea al aire libre, parece siempre a quién lo oye bajo el agua, desagradable, violento y grave”[12]. Por eso se pescan los delfines rodeándolos con barcas y haciendo ruido en el agua, impeliéndoles a huir hacia la playa, donde embarrancan[13]. Y, por eso, los pescadores evitan hacer ruido con los remos y las redes y permanecen callados y conducen a los peces a las redes en un momento dado, con gritos y con piedras, que lanzan en el agua o, como en el caso de la pesca fluvial de los gobios o cotos (κόττους), golpeando con cantos rodados las piedras bajo las que se ocultan, para que salgan[14].
Para Aristóteles, los peces que tendrían el oído más fino serían el mújol o lisa (κεστρεύς), el robalo (λάβραξ)[15], la salpa (σάλπη), “y todos los peces del mismo género”.
Junto con la vista, considera Aristóteles a la audición con el sentido más útil, porque:
“Cada uno a su manera, secundan la vida del pensamiento; el oído posee un valor particular, porque la palabra es el principal instrumento para enseñar y aprender”[16]
III.2. Teoría moderna del sonido.
El sonido se produce por la vibración molecular de un sólido, un líquido o un gas a consecuencia de una presión mecánica producida por eventos tales como un choque, una pulsación, un golpe, fricción, etc., que constituya la fuente sonora.
El sonido se transmite como una onda –al modo de las que se forman cuando se lanza una piedra en el agua- como cambios de presión de las moléculas en el medio elástico en el que se producen y, dependiendo de su intensidad, en los que le rodean. Por tanto, dado precisamente su carácter molecular, no se produce en el vacío porque en él no existen esas moléculas.
La ilustración Il IV.3.1 –muestra los efectos de la compresión del sonido en un fluido.

IV.3.1
Transmisión del sonido en un fluido. Se produce una onda de
presión por compresión, que hace que el resto de las partículas
se compriman entre ellas. Fuente: Wikipedia Commons.
La Il IV.3.2 muestra esquemáticamente la propagación del sonido y su captación por el oído.
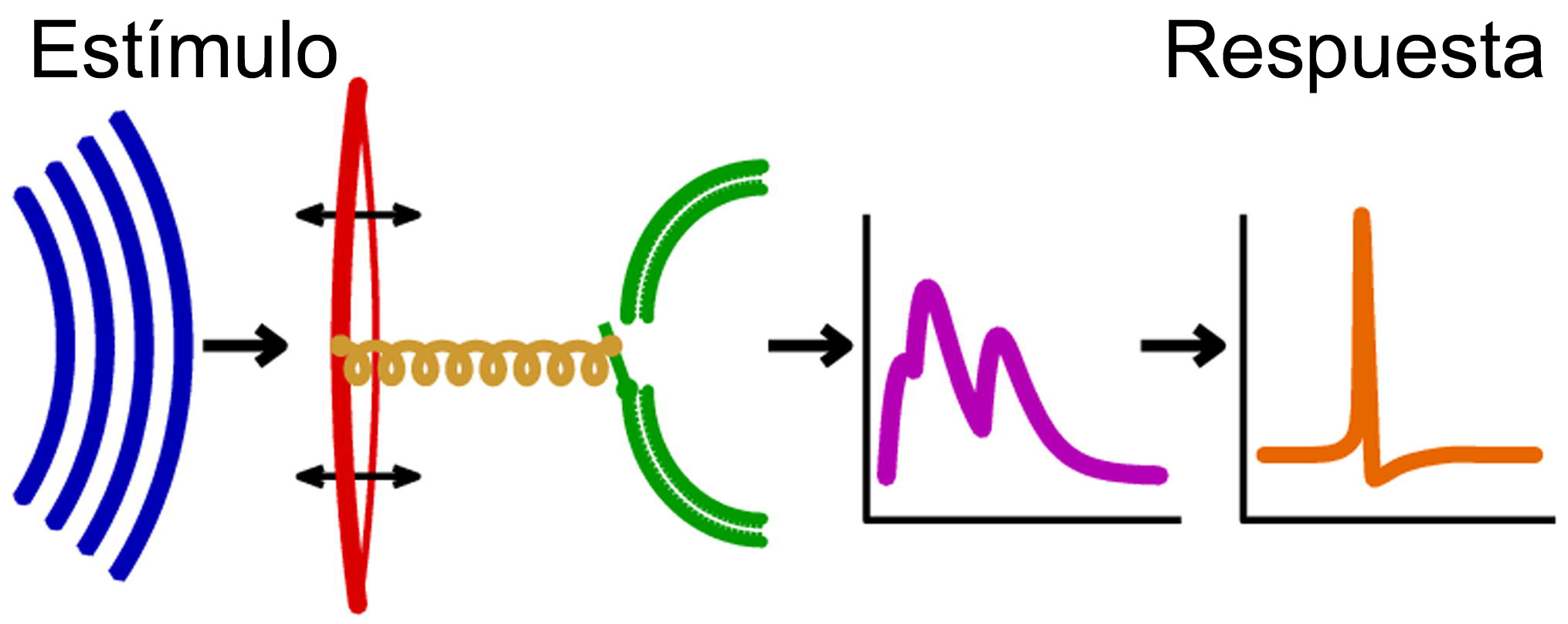
IV.3.2
Azul: Ondas sonoras. Rojo: tímpano. Amarillo: cóclea. Verde: Células
de receptores auditivos. Morado: espectro de frecuencia de respuesta
del oído. Naranja: impulso del nervio. Fuente: Wikipedia Commons.
https://commons.wikimedia.org/wiki/File:Processing_of_sound-es.jpg
Esta onda transmite energía mecánica, producida en su fuente por una presión mecánica, se trasmite, también, como cambios de presión mecánica en el medio que recorre. En los cuerpos sólidos la propagación del sonido se produce específicamente como cambios de la tensión mecánica del cuerpo, expresada en unidades de presión tales como pascales, kg/cm2, etc, que tanto se pueden medir y se pueden percibir por medición directa de la tensión o auditivamente.
Si las vibraciones se producen en la misma dirección hacia la que se propaga el sonido originan una onda longitudinal que produce una percepción aumentada.
La frecuencia de las ondas sonoras es el número de vibraciones completas que se producen cada segundo en la fuente sonora. De forma clara se denominaban ciclos. Hoy, con el propósito de designar las unidades de medida menos populares, con el nombre de un inventor, se denominan Hertzios (Hz) por el apellido de Heinrich Hertz, que probó en la práctica la existencia de las ondas radioeléctricas mediante un oscilador elemental.
El oído humano percibe las frecuencias entre 20 Hz y 20.000 Hz (20 Khz).
Como cualquiera que haya utilizado un silbato para perros sabe, muchos animales perciben sonidos de superior frecuencia a los que nosotros oímos; de forma en consonancia con el aforismo griego de que el hombre es la medida de todas las cosas, se llaman ultrasonidos.
El oído, como el micrófono, convierten la presión mecánica que viaja por el medio –normalmente el aire- en una señal eléctrica. En el caso de los micrófonos más sencillos mediante un condensador colocado entre dos paredes elásticas; y en el oído, como veremos luego.
En todos los sonidos que percibimos, se pueden distinguir tres cualidades: a) Intensidad, que relaciona volumen y potencia acústica. b) Tono relacionado con la frecuencia; y c) Conformación electrógrafica que muestra gráficamente en relación con el tiempo, la intensidad y el tono.
La intensidad de un sonido viene determinada por la amplitud de las vibraciones de la fuente sonora y su frecuencia: puesto que el nivel de potencia acústica es la cantidad de energía radiada al medio en forma de ondas por unidad de tiempo. La amplitud de la señal obedece a la energía instantánea de la fuente sonora; corresponde, pues, a la amplitud de la vibración de la fuente sonora. La frecuencia es el número de veces que la señal se repite durante esa unidad de tiempo, el número de esas vibraciones; de forma que, cuanto menor es la longitud de onda –y por tanto mayor la frecuencia- mayor es la cantidad de energía producida por la onda.
En el caso de una onda sónica esférica que se trasmite desde una fuente puntual en el espacio libre –esto es: sin obstáculos- la intensidad acústica es inversamente proporcional al cuadrado de la distancia entre la fuente sonora y el punto de percepción, por la razón de que la intensidad acústica es inversamente proporcional al área del frente de onda (I=P/4πr2).
La unidad utilizada para medir la potencia es el vatio por metro cuadrado (W/m2) medido en posición normal –esto es: perpendicular- al radio de la onda.
El oído humano tiene la capacidad de oír sonidos a partir de una intensidad de 10-12 W/m2. Esta intensidad se conoce como umbral de audición.
Dado que hay grandes diferencias en el rango de intensidades que el oído humano puede oír sin dolor, para medir la intensidad del sonido percibido se emplea una escala logarítmica, en la que se emplea como nivel de referencia el umbral de audición, y que expresa esa intensidad en decibelios, según la siguiente formula. BdB = 10 log10 I/I0. Donde BdB es el nivel de intensidad acústica en decibelios, I es la intensidad acústica en W/m2 e I0 es el umbral de audición.
En resumen, la intensidad de percepción de un sonido depende del área y la energía de vibración de la fuente sonora y de la distancia a esa fuente; pues la energía vibratoria emitida por la fuente se distribuye uniformemente en ondas esféricas cuya superficie aumenta proporcionalmente al cuadrado de sus radios. La energía que recibe el oído es, por consiguiente, una fracción de la energía total emitida por la fuente; tanto menor cuanto más alejado está el oído. Esta intensidad disminuye 6 dB cada vez que se duplica la distancia a la fuente sonora. Para evitar este debilitamiento se canalizan las ondas por medio de un “tubo acústico” –bocina- y se aumenta la superficie receptora aplicando al oído una trompeta o trompetilla o un amplificador electrónico. La frecuencia produce tonos más agudos cuanto mayor es; esto es: cuanto mayor es el número de vibraciones por segundo de la fuente sonora. La ilustración Il IV.3.3 muestra la representación gráfica de esas frecuencias.
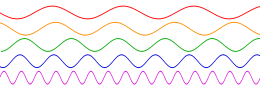
IV.3.3
onda sinusoidal; Variación de frecuencia; Abajo podemos ver las
frecuencias más altas. El eje horizontal representa el tiempo.
Fuente: Wikipedia Commons.
Se llaman sonidos simples aquellos que tienen un solo armónico; caso típico del silbido. No existe, en este caso, ni resonancias, ni vibración de las cuerdas vocales; el sonido se produce, simplemente, por la expulsión del aire por los labios y por tanto, se modifica por la apertura de los labios, simplemente; no tiene más complicaciones.
Frente a estos sonidos simples, observamos sonidos complejos, en los cuales el sonido producido por la fuente sufre resonancias. La resonancia es el fenómeno que se produce cuando dos cuerpos adquieren la misma frecuencia de vibración, porque uno empieza a vibrar al recibir las ondas sonoras emitidas por el otro, como sucede con la guitarra, que se afina y se dota de una caja al objeto de producir esas resonancias, como muchos otros instrumentos musicales.
La señal eléctrica del micrófono puede observarse, como es evidente, mediante un osciloscopio, espectografo o aparato similar, convertida en una señal gráfica temporal; en altura suele representarse la intensidad de la señal en milivoltios y, en longitud, la frecuencia de la señal en milisegundos. El número de oscilaciones por unidad de tiempo, representa la frecuencia.
El sonido simple se representa por una sinusoide. El sonido complejo puede descomponerse en sinusoides, pero el aparato la representa simplemente en el dominio de la frecuencia; es decir, el aparato no distingue más que la frecuencia, y por tanto, muestra un espectro continuo, con el perfil complejo representado por los componentes de ese sonido, y no las líneas diferentes que podrían dibujarse explicativamente, si quisiéramos mostrar la composición del sonido complejo. La señal temporal, no analizada, de las cinco vocales castellanas pronunciadas por un varón, se muestra en la Il IV.3.4
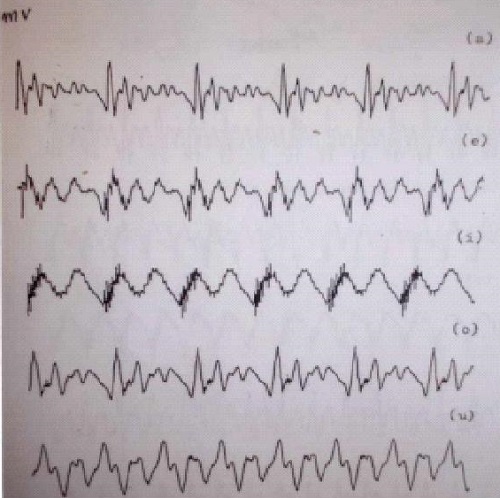
IV.3.4
Fuente: Proporcionada por el Catedrático de Física-Médica
D. José Luís Ramón.
Un armónico es la representación en frecuencia de cada uno de esos sonidos que integran el sonido complejo, representados por separado analizando la imagen que da el aparato.
La transformación de Fourier –positiva e inversa- permite matemáticamente transformar la señal temporal en armónicos o componentes de frecuencias diferentes y, viceversa, en lo que se denomina análisis armónico.
La Il IV.3.5 muestra la transformación de la señal temporal (en rojo) en sus armónicos (en azul) por orden de frecuencia de menor a mayor.
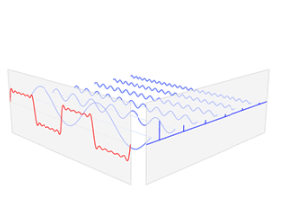
IV.3.5
La transformada de Fourier relaciona una función en el dominio del
tiempo, mostrada en rojo, con una función en el dominio de
la frecuencia, mostrado en azul. Las frecuencias componentes, extendidas
para todo el espectro de frecuencia, son representadas como picos en el
dominio de la frecuencia. Fuente: Wikipedia Commons.
Esta descomposición, que muestra gráficamente la longitud de onda de dos armónicos, su frecuencia, su amplitud –relacionada con el volumen y potencia acústica- y fase, simplifica el estudio de los sonidos complejos, ya que permite estudiar cada armónico independientemente y combinar los resultados por superposición.
En general, cuanto mayor sea la compresibilidad (1/K) del medio conductor de la onda sónica, tanto menor es la velocidad de propagación del sonido. Por guardar relación con la compresibilidad, cuanto mayor sea la densidad del medio conductor –esto es: el peso medio del material que constituye el medio por metro cúbico (ρ) tanto mayor será la velocidad de propagación del sonido. Por tanto, la velocidad de propagación del sonido se relaciona con las dos magnitudes citadas: 1/K y ρ .
La velocidad de propagación del sonido depende, pues, de las características del medio en que se realiza dicha propagación y no de las características de la onda o de la fuerza mecánica que la genera[17].
En los sólidos, la velocidad de propagación del sonido es superior a la que se produce en los líquidos y los gases. Y esa velocidad es particularmente superior en los metales, dado su particular enlace molecular a través de las bandas de valencia y las bandas de conducción de sus electrones (enlace metálico) y su estructura cristalina en estado sólido.
En los sólidos, la velocidad del sonido resulta de la formula Vs = √E/ρ
En la que E es el módulo de Young o módulo de elasticidad longitudinal, parámetro que expresa porcentualmente la elongación de un material hasta su límite elástico, determinado empíricamente mediante un ensayo de tracción. Ya hemos señalado que ρ representa el peso medio del material que constituye el medio por el metro cúbico.
En la madera, la velocidad de propagación del sonido es 3.700 metros por segundo (3.700 m/s). En el hormigón, 4.000 m/s. En el cobre, 4.650 m/s. En el vidrio, 5.260 m/s. En el hierro, 5.930 m/s. En el aluminio, 6.400 m/s.
En los líquidos, la velocidad del sonido se expresa por la siguiente formula: Vs=√K/ρ
En la que K es el módulo de comprensibilidad que mide la resistencia del líquido a la comprensión uniforme y, por tanto, indica en pascales –los antiguos bares- el aumento de presión requerido, para causar una disminución del volumen del líquido en un 5%. Evidentemente, es el equivalente al módulo de Young que se utilizaba en la formula anterior para medir la elasticidad de un sólido. ρ (ro), corresponde a la densidad, como en la formula anterior.
La velocidad de propagación del sonido en el aceite de linaza es de 1.770 m/s. En el agua destilada, a 20º, 1.482 m/s. En el mercurio líquido, 1.451 m/s. En el aceite de oliva, 1.430 m/s. Y en el alcohol metilico, 1.130 m/s.
En los gases, la velocidad del sonido la expresa la ecuación siguiente: V=√γ RT/M.
En la que γ (gamma) es el coeficiente de dilatación adiabática del gas, o relación entre el calor específico de un gas a presión constante y el calor específico de ese mismo gas a volumen constante y en ambos casos adiabáticamente; esto es: sin intercambio de calor entre el gas en cuestión y el medio que lo rodea.
El aire que se calienta equivale, termodinámicamente, a aire que gana energía, y el que se enfría, a aire que la pierde. El calor específico es el número de calorías necesarias para elevar la temperatura de un gramo de cualquier sustancia en un grado centígrado.
Siendo en realidad una función de la temperatura, el calor específico variará según la temperatura inicial al que estuviese la sustancia. Para el agua, es una caloría exactamente cuando se eleva su temperatura de 14,5 a 15,5ºC. En la práctica meteorológica pueden sin embargo considerarse los calores específicos como muy aproximadamente constantes; ya que los coeficientes térmicos de dilatación de un gas a presión constante (Cp) y de aumento de presión a volumen constante (Cv), son iguales entre sí y tienen el mismo valor para todos los gases perfectos (Ley de Gay Lussac y Boyle y Mariotte).
Un gas perfecto es aquel que tiene completa movilidad, elasticidad, expansibilidad y comprensibilidad ilimitada y que, de acuerdo con las leyes de Mariotte y de Joule, a una temperatura constante, cualquiera que sea, posee una presión inversamente proporcional a su volumen específico y una energía independiente de su volumen.
Aún cuando la sustancia en cuestión sea un gas, como ocurre en el caso del aire, caben infinitas evoluciones posibles en el proceso de su calentamiento y, por tanto, la posibilidad de infinitos calores específicos; pero a los efectos de la ecuación que estamos analizando caben solamente dos: el calor específico o volumen constante (Cv) y el calor específico a presión constante (Cp).
Como ya he dicho, el calor específico se define como el número de calorías que hay que comunicar a un gramo de aire seco para elevar en un grado centígrado su temperatura, manteniendo invariable su volumen en el primer caso, y permitiendo libremente su dilatación, en el segundo. Una imagen tangible de ambos, nos la daría el proceso de calentamiento de un gramo de aire encerrado, respectivamente, en una bombona metálica indilatable y en un globo de goma perfectamente elástico.
Los valores respectivos, para el caso del aire atmosférico son Cv=0,17 y Cp= 0,24 calorías/gramo.
γ (gamma) es el resultado de dividir Cp por Cv; o sea, Cp/Cv que equivale a 1,4.
El aire seco a 20ºC tiene pues un coeficiente o cociente de dilatación adiabática de 1,40, como el deuterio (H2) que tiene 1,41, como el oxigeno común o biatómico (O2), o como el oxido de carbono (CO) a la misma temperatura.
R, en la ecuación analizada, es la constante de gas universal o constante absoluta de los gases, que relaciona los valores de presión, volumen y temperatura absoluta de un gas, teniendo en cuenta que el calor que se comunica a un gas se emplea en aumentar su energía interna (Cv x T) y a la vez en el trabajo de su expansión (p x v, siendo p la presión y v el volumen), según el primer principio de termodinámica.
La constante expresa en julios o joules –unidad derivada del Sistema Internacional utilizada para medir energía y trabajo (equivalente a un vatio/segundo)- y calor (equivalente a 0,238 calorías)
Como, según la Ley de Avogadro-Ampere, a igualdad de volumen, presión y temperatura, todos los gases tienen el mismo número de moléculas, la constante expresa la relación de la cantidad de materia en un gas ideal, expresada en número de moléculas gramo o moles[18], con la presión en bares o pascales, el volumen (v) en cm3 y la temperatura (T) en grados Kelvin, o temperatura absoluta, en la que 0º es equivalente a -273ºC-: la temperatura del hidrogeno líquido.
Por lo que la mencionada constante tiene el valor universal de 8,314 J/mol K o 8,314 kg.m2 /mol.k.s2 que corresponde a la llamada ecuación de estado de los gases perfectos; pues según la Ley de Avogadro-Ampere, a igualdad de volumen, presión y temperatura todos los gases tienen el mismo número de moléculas y, por tanto, la constante R no depende más que de la masa del gas en cuestión, determinada por la suma de la masa atómica de los elementos que componen sus moléculas.
En la ecuación que estamos utilizando, expresiva de la velocidad del sonido en los gases, T es la temperatura en grados absolutos o grados Kelvin. En los gases, como en todos los medios en los que se transmite el sonido, la temperatura aumenta la movilidad molecular y, con ella, la velocidad del sonido; pero en los gases es un factor de la mayor importancia. En el caso del aire atmosférico seco es conocido que cada grado centígrado de aumento de su temperatura provoca un aumento de 0,6 m/s de su velocidad. M, por último, expresa en la ecuación la masa del gas en cuestión, expresada en el peso en kilogramos de una molécula gramo o mol del gas, que como ya hemos dicho en los gases perfectos equivale a unos 22,4 L de ese gas y que se puede calcular, aproximadamente[19], sumando las masas moleculares de los átomos constituyentes del gas en cuestión.
De todo lo expuesto, es posible establecer ciertos criterios generales sobre la velocidad del sonido en los distintos medios:
1) La velocidad de propagación de la onda sonora depende de las características del medio en que se realiza y no de las características de la onda o de la energía que la genera.
2) En general, cuanto mayor sea la compresibilidad del medio (1/K) tanto menor es la velocidad del sonido que a través de él se propaga y cuanto mayor sea la densidad del medio, tanto mayor es la velocidad de propagación del sonido.
3) Por tanto, la velocidad del sonido, con carácter general, relaciona ambas magnitudes mediante la fórmula: V= √K/ρ.
Como hemos expuesto antes, esa formula varia entre los medios sólido, líquido y gaseoso y precisamente para adaptarse a la forma de medir en cada medio de la forma más eficaz K y ρ.
4) Por ello, en general, el sonido se propaga más rápidamente en los sólidos que en los líquidos y en los líquidos más rápido que en los gases.
5) Como en los gases la temperatura influye tanto la compresibilidad como la densidad, suele ser la temperatura el factor de mayor importancia.
III.3 Absorción, reflexión, reverberación y resonancia.
Las ondas concéntricas y circulares que provocamos lanzando una piedra al agua, los rayos de luz que inciden sobre un espejo o sobre una superficie que no absorbe completamente la luz y hasta las ondas radioeléctricas que inciden sobre las superficies metálicas sufren alteraciones en su dirección, siguiendo modelos muy parecidos.
En principio, toda onda que choca contra un cuerpo produce cuatro efectos:
1) Por el de absorción, ve disminuida su energía y pierde alcance y e intensidad.
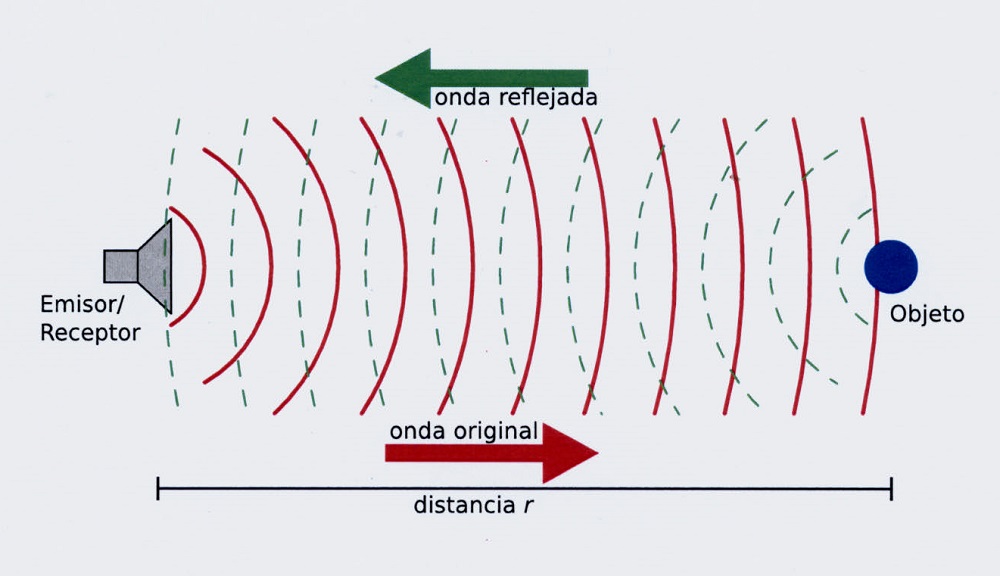
2) Por el de reflexión, rebota contra el obstáculo, que devuelve al medio de transmisión una onda de características iguales pero de menor energía.
3) Por el de difracción, la onda cambia de dirección, y eventualmente de forma como consecuencia del choque contra un obstáculo material.
4) Por el de refracción, la onda cambia de dirección debido a las variaciones del medio transmisor.
La ilustración Il IV.3.6 esquematiza todos estos efectos
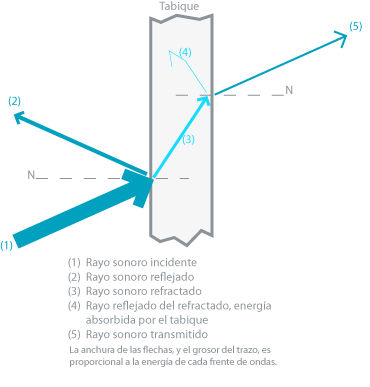
IV.3.6
Fuente: Creative Commons
Cuando una onda sonora incide sobre una superficie plana, como un tabique, se produce en primer lugar una absorción, cuya magnitud depende del material de la misma; porque aunque todos los materiales absorben, no lo hacen en la misma proporción: el algodón absorbe el 86% de la energía de la onda de un 1Khz, la madera el 70%, mientras el cristal absorbe el 58% de la energía de la señal.
El concepto de coeficiente de absorción fue desarrollado por Wallace Sabine que estableció que una ventana abierta que no refleja ningún sonido actual, representa el absorbente perfecto y le asigno un coeficiente de absorción de 1 –esto es: del 100%-. En otros obstáculos hay siempre reflexión y refracción parciales. Una parte de la energía se absorbe y otra se refleja o refracta por el obstáculo.
El segundo efecto que numeramos es el de reflexión. La energía de la onda no absorbida se refleja de dos formas distintas: a) si la superficie del obstáculo no es lisa, las ondas de mayor intensidad que son las que coinciden con el ángulo incidente, serán reflejadas en todas direcciones, -según el principio de Huygens- en reflexión irregular o difusa.
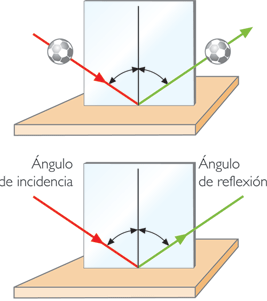
Si la superficie del obstáculo es lisa, el resto de la energía de la onda no absorbida se refleja según las leyes de reflexión de las ondas: 1) Todos los puntos que se enfrentan a la onda sonora se convierten en otros tantos centros reflectores, emisores de un sistema de ondas llamadas de “retroceso o reflexión” de la misma frecuencia y amplitud. 2) El ángulo de incidencia en relación a la perpendicular de la superficie reflectante, es igual al de reflexión, como se observa en las Il IV.3.7 y Il IV.3.8. 3) En el mismo medio, la onda reflejada tiene la misma velocidad que la incidente; pero como hemos dicho, no la misma energía en virtud de la absorción.
IV.3.7
Fuente: Wikipedia Commons
IV.3.8
Fuente: es.slideshare.net
A causa de la reflexión se oye el eco producido por una superficie plana grande –mayor que la longitud de onda que debe reflejar- perpendicular a la propagación de la onda sonora y distante entre 17 y 34 metros, según el tiempo de persistencia el tipo de sonido, para que pueda distinguirse el sonido reflejado por esa superficie del sonido producido por la fuente sonora.
Por el efecto de reflexión se canaliza la onda sonora para mejorar su percepción evitando la dispersión esférica de la energía y canalizando la onda en el sentido longitudinal en el que se produce, mediante tubos acústicos, tornavoces, placas de reflexión, techos abovedados, conchas de apuntador, –que proyectan la onda sonora con precisión desde un punto del foco de la elipse, donde esta la fuente sonora, hasta otro, donde se encuentra el auditorio, -evitando la obstaculización, por difracción, producida por los obstáculos que se hayan en la línea visual directa, como las cabezas de los espectadores.
En el ambiente natural, son superficies de reflexión potenciadoras del sonido el suelo, los precipicios y acantilados, las nubes –que amplifican y prolongan el sonido del trueno y del cañón- y las capas profundas de la nieve, de temperatura más baja, donde se refleja más eficientemente el sonido.
Se produce reverberación cuando las ondas reflejadas llegan al oyente antes de que se extinga la sensación auditiva que produce la onda directa, por el efecto de persistencia acústica cerebral del sonido que transmite la onda directa, que es de 0,1 segundos para sonidos musicales y de 0,07 segundos para la palabra (efecto Haas o de precedencia). En este último caso, el agente percibe la onda directa, y sin solución de continuidad, como “cola sonora” las sucesivas reflexiones que enfatizan los sonidos musicales y frecuentemente –por su corto tiempo de persistencia acústica, dificulta entender las palabras. Se define como tiempo de reverberación el que transcurre hasta que la intensidad del sonido queda reducida a una millonésima de su valor inicial.
La reflexión no solo se produce sobre cuerpos sólidos y sobre gases o fluidos de diferente naturaleza química. Un mismo sólido o fluido a diferentes temperaturas la produce también. Por eso, reflejan las nubes el sonido del cañón y el trueno; por eso refleja el sonido potenciando su alcance, la reflexión sobre las capas de nieve más profundas y frías, o disminuyen ese alcance, en los días acústicamente opacos, las columnas y capas de aire a temperaturas diferentes, de forma que las ondas sonoras son parcialmente reflejadas en cada paso por aire a diferentes temperaturas, a causa de que, como la velocidad de propagación de las ondas sonoras varia con la temperatura del medio de transmisión y con la humedad relativa del aire, cuando una onda pasa en un medio en la que se mueve a distinta velocidad, o a otro medio, como vimos al principio, es parcialmente reflejada en la superficie de contacto entre las dos zonas o los dos medios.
Es resonador todo cuerpo capaz de vibrar reforzando el sonido de una tonalidad determinada. Disponiendo de una serie de resonadores de frecuencias de resonancia conocidas puede verificarse fácilmente el análisis de la tonalidad de los sonidos y reforzar los sonidos que emiten los instrumentos musicales.
III.4 Difracción del sonido.
La difracción es un fenómeno característico de las ondas que, por tanto, ocurre tanto en las ondas electromagnéticas –la luz y las ondas de radio-, o en las ondas en la superficie de un fluido, como en las ondas sonoras.
Cuando una onda sonora incide perpendicularmente sobre una superficie plana y lisa más grande que su longitud de onda –y esta longitud de onda puede ser hasta de 20 metros en la gama de sonidos audibles por el oído humano- se refleja como una onda esférica que tuviera su centro en un punto, perpendicular a la superficie reflectante, a igual distancia de la superficie reflectante, pero detrás de ella, que la distancia a la que se encuentra delante de la superficie reflectante la fuente sonora.
La Il IV.3.9 muestra gráficamente este efecto que responde a un fenómeno prototípico de reflexión.
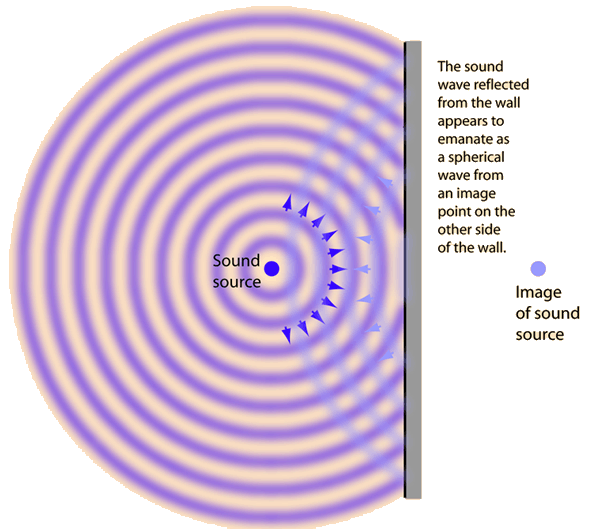
IV.3.9
Por el contrario, cuando la superficie reflectante no es lisa, o no es plana, o es de tamaño inferior a la longitud de la onda sonora, o muestra aberturas, pasos o disrupciones superiores a la longitud de la onda sonora, se produce la difracción, en la que -según el principio de Huygens- todos los puntos de un frente de ondas inicial que son alcanzados en el mismo momento por un movimiento ondulatorio, deben ser considerados como centros productores de un nuevo sistema de ondas esféricas secundarias que se extienden en todas las direcciones con la misma velocidad, frecuencia y longitud de onda que el frente de onda de que procede.
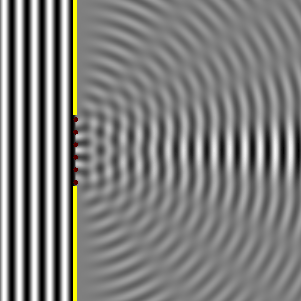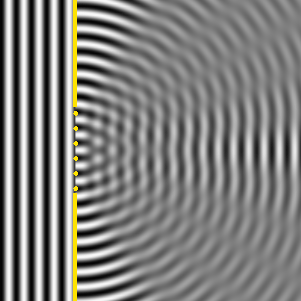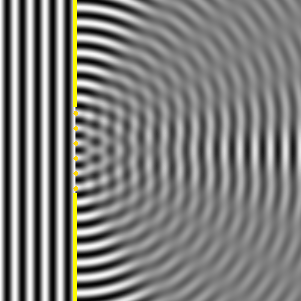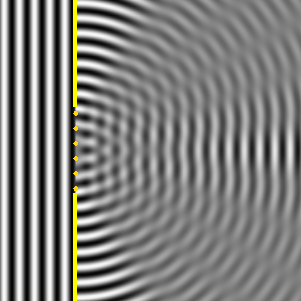
Ese nuevo frente de onda resultante, son las ondas difractadas. Cuando el tamaño de la rendija que obstaculiza el paso de la onda es igual al de la longitud de onda, la difracción es total o perfecta, y la intensidad recibida detrás de la rendija disminuye lentamente desde el máximo situado en frente de ella al mínimo situado a sus lados, como se observa en la Il IV.3.10. Por el contrario, cuando la rendija es cinco veces más grande que la longitud de onda, la difracción se produce cerca de los bordes de la rendija, la máxima intensidad se produce también detrás del centro de la apertura, pero la intensidad de la señal decrece rápidamente y la difracción solo se observa claramente en la proximidad de los bordes de la abertura y detrás del obstáculo; además las ondas difractadas en las proximidades de cada borde, se amortiguan y por ello la intensidad decae bruscamente, así se observa en la Il IV.3.11.

IV.3.10
Difracción de una onda plana cuando el ancho de la ranura
es igual a la longitud de onda.
Fuente: Wikipedia Commons
IV.3.11
Difracción de una onda plana cuando el ancho de la ranura
es varias veces la longitud de onda.
Fuente: Wikipedia Commons
Las ondas de alta y baja frecuencia no se reflejan de igual forma. Como la longitud de onda de las bajas frecuencias (20 Hz) en el medio aéreo es de unos 17 metros, esas ondas son capaces de rodear la mayor parte de los obstáculos; las de alta frecuencia (20 Khz) tienen por razón de su frecuencia y de la velocidad del sonido, una longitud mil veces menor -17mm- y, por tal razón no rodean los obstáculos, por lo que se producen “sombras acústicas” detrás de ellos y reflexiones o rebotes sobres sus superficies expuestas a la onda directa.
Por razón de la amortiguación de la señal sonora que produce la difracción, se puede afirmar que cuando se preserva la línea de visión a un escenario, no sólo se mejora la percepción visual, sino también la auditiva que, de no emplearse superficies reflectantes eficientes, sufre la difracción producida por las cabezas de los espectadores.
III.5 Refracción del sonido
Como vimos al principio, en la superficie de separación de dos medios aptos para conducir el sonido o de dos zonas del mismo medio, con diferente velocidad de transmisión, además de producirse un fenómeno de reflexión, la onda traspasara la superficie de separación, trasmitiéndose tras esa superficie al segundo medio con un cambio de dirección de la propagación de la onda cuando no ha incidido perpendicularmente sobre la superficie de separación, según las siguientes leyes de Descartes/Snell:
1ª. El rayo incidente[20] y el refractado están en un mismo plano; son coplanarios.
2ª. La relación entre el seno –esto es: la función angular- del ángulo de incidencia y el seno del ángulo de refracción, es igual al cociente de dividir la velocidad de propagación del sonido en el primer medio –o en la primera zona de distinta velocidad de propagación, en el mismo medio- por la velocidad de propagación del sonido en el segundo medio, o en la segunda zona de distinta velocidad de propagación del mismo medio, como se ilustra gráficamente en la Il IV.3.12; en la que A-B es la superficie de separación, siendo menor la velocidad de propagación del segundo medio o de la segunda zona de distinta velocidad de propagación figurada en la parte inferior de la ilustración, I-o es una onda incidente en o ó cero que forma con la perpendicular a la superficie de separación un ángulo i, y tras incidir en la superficie de separación se refracta en la dirección o ó 0-R formando un ángulo r con la perpendicular a la superficie de separación, menor que el ángulo i por las razones y en virtud de la Ley 2ª de Descartes/Snell. El seno del ángulo i es el resultante de dividir el cateto opuesto al ángulo i (I-M) por la hipotenusa (I-o); o sea:
sen i=IM/Io
Igualmente:
sen r= RM´/Ro
Pero, como Io es igual a Ro, por ser ambos radios de la misma circunferencia, resulta:
sen i/sen r = IM/RM
Si se aumenta o disminuye el ángulo de incidencia i aumentará o disminuirá, también r, en la misma proporción de la relación entre los senos de ambos ángulos, siempre que i no llegue a alcanzar 90º y resulte, por tanto, perpendicular a la superficie de separación, en cuyo caso no se produce difracción.
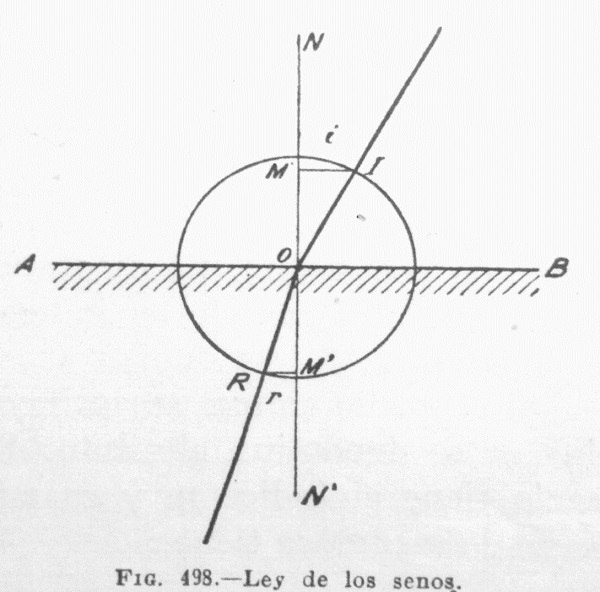
IV.3.12
El cambio de presión de mayor influencia en la práctica en la transmisión aérea del sonido es el debido al viento. La velocidad del viento aumenta prácticamente siempre con la altitud y es de particular importancia en las capas de contacto con la tierra –por eso, a efectos meteorológicos, la velocidad del viento se mide a una altitud mínima de 10 metros sobre la tierra, porque debajo de ella, esta demasiado disminuida por la fricción contra la superficie terrestre- provocando que la onda sonora que se dirige en sentido contrario al viento se desvíe hacia arriba al sufrir la onda esférica presión adversa y sobre capas en las que el viento es cada vez más rápido; mientras que la onda sonora a favor del viento ve favorecida su difusión por la presión y la reflexión sobre el suelo, como se observa gráficamente en la Il. IV.3.13.

IV.3.13
Como vimos al hablar de la velocidad del sonido en los gases, la temperatura aumenta la velocidad del sonido. Por tanto, en las horas diurnas, cuando el aire en contacto con el suelo esta más caliente, el sonido se desvía en virtud de la refracción y del ascenso del aire caliente hacia las capas más altas; independientemente de la reflexión y debilitamiento que esas columnas de aire producen en las ondas sonoras. Por tal razón, la percepción de la onda sonora se dificulta. La Il IV.3.14 muestra gráficamente ese efecto.
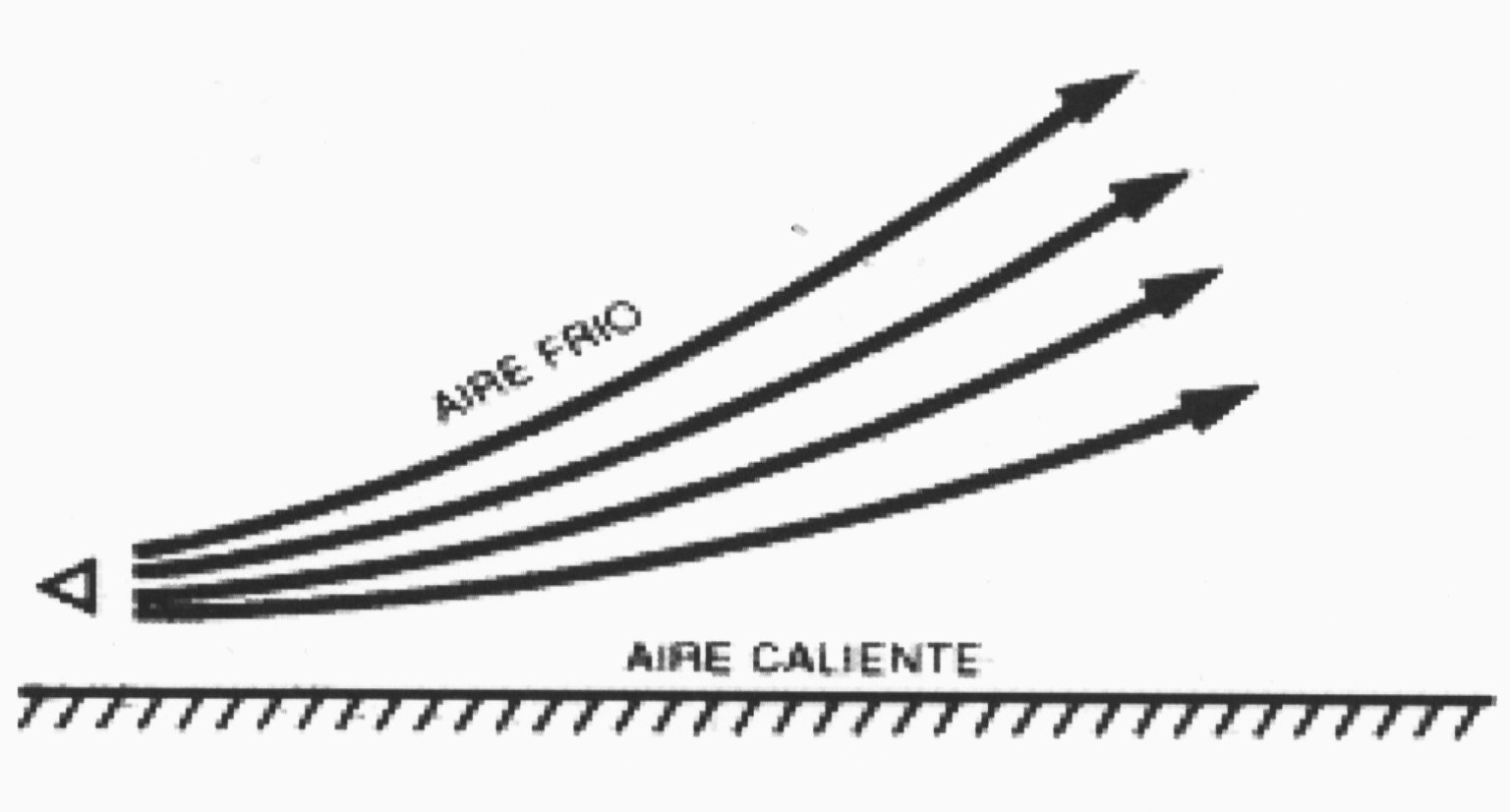
IV.3.14
Por el contrario, durante las horas nocturnas, el rápido enfriamiento de la superficie de la tierra hace que el aire frío –salvo fenómeno meteorológico particular- se encuentre en contacto con ella y, en ese caso, por las razones contrarias, las ondas sonoras se propagan horizontalmente y se reflejan en el suelo prolongando su alcance -Il. IV.3.15- que es el mismo efecto que puede producirse en tiempo muy frío cuando las ondas sonoras rebotan sobre las capas más frías de la nieve profunda.
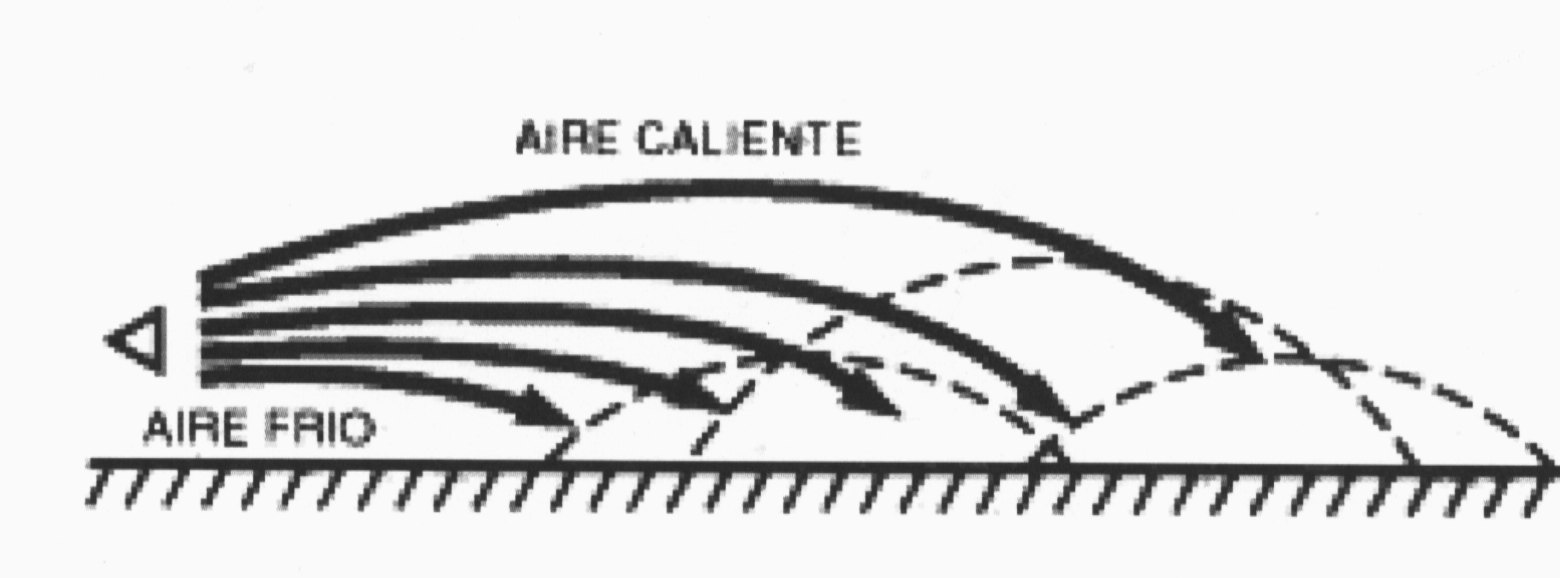
IV.3.15
III.6 Fisiología de los mamíferos.
Ya hemos visto que el sonido es una onda causada por una presión mecánica y que la onda sonora transmite longitudinalmente los efectos vibratorios causados por esa presión mecánica. Se origina, pues, por una presión y como consecuencia, el organismo la percibe también como una presión, por medio de un tipo específico de mecanosensores.
El esquema del mecanosensor que percibe la señal sonora es, pues, de los mas sencillos: un canal iónico celular que, bajo la presión de la onda sonora se abre, permitiendo la entrada en la célula de iones positivos de sodio (Na+), como vemos en la Il IV.3.16, en un proceso que describiré en detalle un poco más tarde.
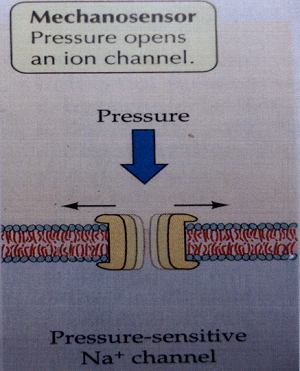
IV.3.16
Fuente: Life, The Science of Biology.
Se trata, pues, de una distorsión física de la membrana plasmática de esas células especializadas neuronales que causa la apertura de esos canales iónicos y altera su potencial eléctrico, produciendo una potencia eléctrica de acción como la que vimos en los fotosensores del sentido de la vista.
Otro tipo de mecanosensores que no son neuronales son las células ciliares –esto es: en forma de pelo- llamadas stereocilias que tienen en conjunto el aspecto de tubos de órgano. Cuando estas stereocilias se doblan, alteran proteínas receptoras en su membrana plasmática. Cuando se doblan en una dirección su carga eléctrica se vuelve más negativa y cuando se doblan en la dirección opuesta más positiva. Cuando su carga eléctrica se vuelve más positiva, estas células ciliares liberan neurotransmisores a las neuronas sensoriales asociadas a ellas y éstas envían señales eléctricas al cerebro.
Los órganos humanos de audición, por su relativa complejidad debida a la evolución, permiten hacerse una idea bastante completa anatómica y funcional del sistema.
Las orejas son los apéndices auditivos que reciben en sus pabellones las ondas sonoras y las dirigen al canal auditivo que conduce a la percepción, en el oído medio y en el oído interno. Si se observa como los conejos y los caballos cambian la orientación de sus orejas para dirigir su atención a una fuente de sonido en particular, se comprende bien a las claras la función de estos apéndices.
La estructura del oído humano se representa en la ilustración IV.3.17 mediante cinco diagramas, en orden de magnificación progresiva[21].
Como se ve del primero de estos diagramas, la membrana del tímpano cubre el final del canal auditivo y vibra en respuesta a las ondas de presión recibidas en el pabellón auditivo y que recorren su canal, al que nos estamos refiriendo.
En la membrana timpánica, comienza el oído medio, cuya cámara –llena de aire- comunica con la garganta a través del tubo de Eustaquio, equilibrando la presión de la cámara del oído medio con la presión ambiental y reduciendo, cuando se produce una oclusión de dicho tubo, la sensibilidad de la membrana timpánica, como consecuencia del aumento de la presión en la cámara del oído medio.
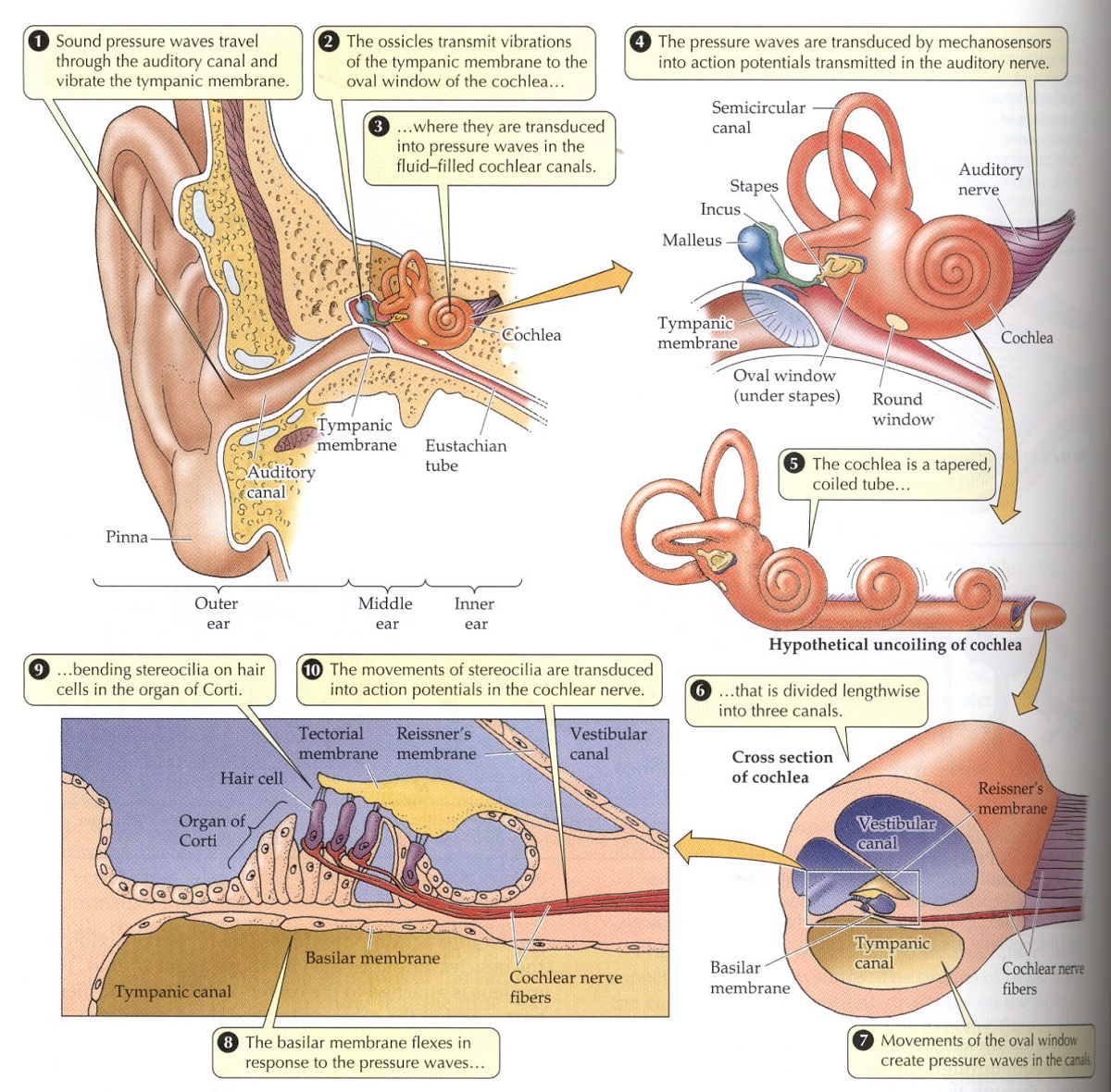
IV.3.17
Fuente: Life, The Science of Biology.Sunderland. Mass. 1997, pag. 938
El oído medio contiene tres delicados huesecillos: “martillo”, “yunque” y “estribo”. Estos huesecillos transmiten las vibraciones de la membrana timpánica a la cóclea, amplificándolas unas 20 veces a causa de su brazo de palanca, a través de la ventana oval de la cóclea, sobre la que se inserta el estribo que, a través de esa ventana, produce cambios de presión en el líquido de la que está llena la cóclea, de forma que en ella se convierten las ondas sonoras de ondas de presión en medio aéreo a ondas de presión en medio líquido.
La cóclea tiene el nombre latino que sugiere su forma -“caracol o concha”- constituye el oído interno, un tubo largo, estrecho y en forma de espiral constituido por tres canales paralelos: vestibular, timpánico y medio o coclear, separados por dos membranas: la basilar y la de Reissner.
Sobre la membrana basilar se encuentra el órgano de Cortí que convierte o traduce las ondas de presión en potencial eléctrico, que a través del nervio auditivo pasa al cerebro. Este órgano contiene los receptores auditivos en forma de pelos o cilios –células claras- cuyos extremos –estereocilios- están en contacto, por el lado opuesto a la membrana basilar, con la membrana rígida tectorial. Cuando la membrana basilar flexiona, como consecuencia de los cambios de presión, los estereocilios flexionan también, al estar apoyados en la membrana tectorial rígida. Como consecuencia de ello, las células claras o células ciliares despolarizan o hiperpolarizan, alterando el potencial eléctrico trasmitido al cerebro desde el ganglio espiral de Corti que es donde se agrupan las neuronas conectadas a las células claras y desde donde parte el nervio auditivo.
Una imagen simplificada, y desenrollada de la cóclea permite explicar como se perciben las diferentes frecuencias sonoras (Il IV.3.18). En ella no se representan más que dos canales separados por la membrana basilar, ya que la función de la membrana de Reissner es contener una específica linfa para el órgano de Corti, separada de la linfa que contiene el resto de la cóclea, función importante para la nutrición del órgano de Corti; pero que no tiene ningún papel en la conversión de las ondas sonoras.
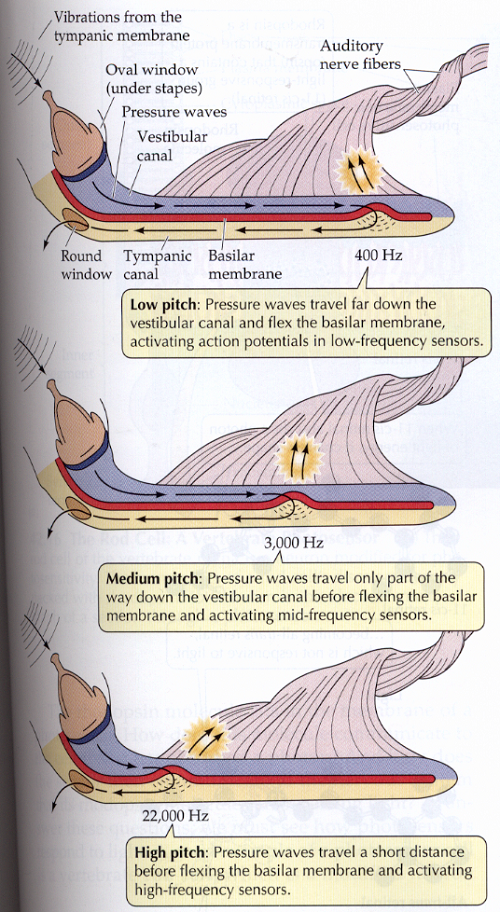
IV.3.18
Fuente: Life, The Science of Biology. Sunderland. Mass. 1997, pag. 939
La capacidad de la membrana basilar para responder a las ondas de presión depende de su estructura. Cerca de las ventanas oval y redonda de la cóclea, en el extremo más próximo al tímpano, la membrana basilar es estrecha y más resistente, pero gradualmente se ensancha y resulta más flexible hacia el extremo opuesto o distal. De forma que la membrana proximal –la que se encuentra en el lado opuesto a la distal- solo oscila bajo la onda de presión más enérgica de altas frecuencias, mientras que la membrana distal flexiona bajo ondas de presión de menor frecuencia, según se observa en la Il IV.3.19. Los potenciales eléctricos que los movimientos de los estereocilios generan son conducidos por medio del nervio coclear al ganglio de Corti y de él al cerebro por medio del nervio auditivo.
Un sonido complejo de muchas frecuencias deforma la membrana basilar en muchos lugares simultaneamente, activando en ellos un conjunto de esterocilios diferentes.
III.7 Fisiología de la audición de los demás animales.
Los peces a los que Aristóteles se refiere, no tienen oído externo ni medio; perciben las vibraciones sonoras de distintas formas.
En primer lugar –en orden, a la evolución natural- a través de la línea lateral, presente en todos los peces y en algunos anfibios, constituida por filamentos nerviosos que recorren el cuerpo del pez en sendas líneas laterales medias que se unen en la cabeza, una a cada lado e incluso otra dorsal, constituidas por células ciliares unidas en forma de cúpula, ligadas a terminaciones nerviosas, mecanoreceptoras representadas en la Il IV.3.19 y que, como ocurría en el oído interno humano, convierten las vibraciones producidas por las corrientes y las vibraciones sonoras de baja frecuencia en impulsos eléctricos. Esa línea permite la percepción de corrientes, cuerpos en movimientos y barreras, ayudando a los cardúmenes de peces a nadar en formación, y funciona como un superoído que recibe pequeñas vibraciones del agua circundante, lo que permite al pez ubicar objetos alejados, así como navegar en aguas oscuras y detectar movimientos de piezas y predadores a distancia.
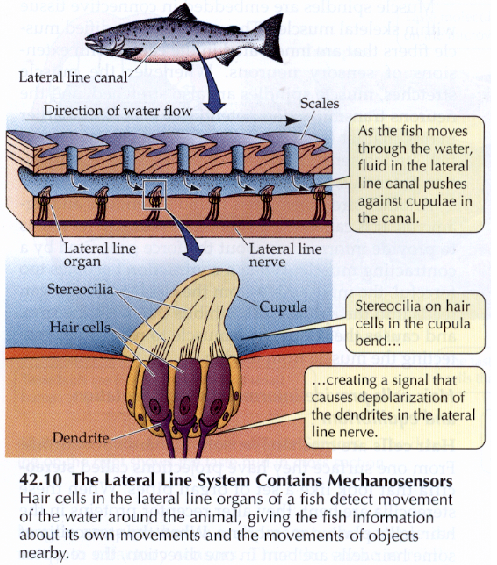
IV.3.19
Fuente: Life, The Science of Biology.Sunderland. Mass. 1997, pag. 936
Perciben también los peces las variaciones de la presión hidrostática como vibraciones a través de la vejiga gaseosa o vejiga natatoria, bolsa localizada en la cavidad celomica -la segunda que aparece en el desarrollo embrionario- del pez, derivada de pulmones ancestrales, cuya pared esta compuesta de fibras de colágeno y es prácticamente impenetrable por el gas, revestida por células sensoriales que convierten los movimientos del aire en impulsos nerviosos y, veremos más tarde, también enlazada con los sonidos que producen los peces.
La función de esta vejiga es, también, la de percibir la profundidad de inversión, durante los desplazamientos del pez; ya que, a medida que cambia de profundidad, la vejiga tiende a dilatarse o comprimirse por el efecto de la presión. A tal efecto, los teleósteos (peces óseos) plesiomórficos mantienen una conexión de la vejiga con el esófago (el ducto neumático) y pueden tomar gas de la superficie para llenar la vejiga. Los teleósteos adultos más evolucionados tienen la vejiga natatoria desconectada del tubo digestivo (fisoclistos) y regulan el volumen de aire de la vejiga mediante una glándula unida a la misma y activada por los jugos hepáticos.
Las vejigas natatorias derivan de pulmones primitivos, faltan o realizan a la vez funciones pulmonares en los peces pulmonados y poliptéridos o bichises[22] y faltan o están reducidas en peces de aguas profundas o abisales. Evidentemente, además de detectar la presión hidroestatica detectan la presión de las ondas sonoras, hasta el punto de que en el superorden de los ostariofisiarios, el segundo orden más grande de los teleósteos -y que comprende especies tan conocidas como los bagres y las carpas, y el 68% de los peces de agua dulce- la vejiga natatoria esta conectada con el laberinto del oído interno a través de unos huesos pequeños derivados de la espina dorsal –tripus, intercalarium, claustrum, scafium- similares a los del oído medio de los mamíferos, en un conjunto anatómico llamado aparato de Weber (Il IV.3.20); pues esos pequeños huesos unidos actúan también a modo de palancas que trasmiten al oído las vibraciones de la vejiga rotatoria.
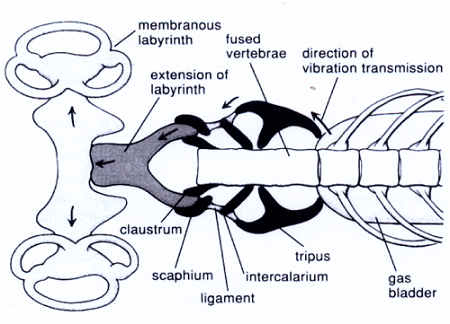
IV.3.20
A estos peces se les suele llamar “peces que escuchan” precisamente por ese rasgo anatómico. El oído interno de los peces se encuentra detrás del ojo, protegido por el cráneo. Consiste morfológicamente en dos partes: una cámara superior o utrículo y una cámara inferior en forma de bolsa o saculo con una pequeña excrecencia inferior llamada Lagena, órgano precursor de la cóclea.
Unidos al utrículo hay tres canales semicirculares llenos de endolinfa que forman ángulos rectos entre sí y que terminan en una pequeña protuberancia o ampolla (Il IV.3.21).
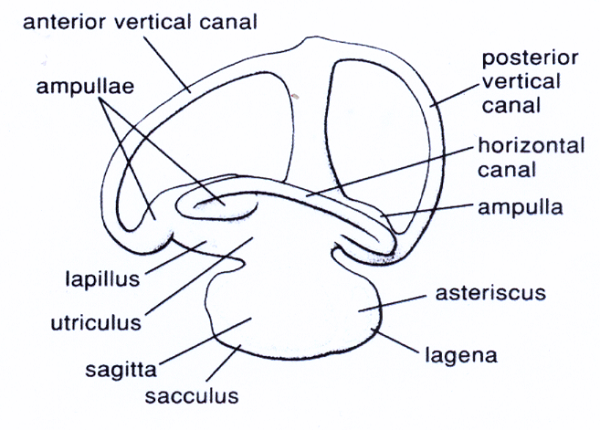
IV.3.21
El saculo y la lagena son los centros de la audición por medio de las células ciliadas y de las terminaciones nerviosas. Las frecuencias que pueden percibir los peces oscilan entre 13 Hz y 17 Khz por segundo, si bien no hay ninguna especie que sea sensible a todas esas frecuencias.
Son los ostariofisios, gracias a su aparato weberiano los que perciben una mayor escala de frecuencias entre 16 Hz a 7 Khz.
Otro método de amplificación aparece en los arenques y boquerones. En ellos la vejiga natatoria tiene prolongaciones que conducen al utrículo del oído interno[23].
Los anfibios que suceden a los peces en la escala filogénetica –es decir que suceden a los peces en la evolución natural- tienen un oído que posee ya una caja de resonancia adecuada; porque la audición en el aire, requiere un sistema más sensible que en el agua, a través de la vejiga natatoria y de los huesos del cráneo.
Durante la metamorfosis, el renacuajo posee línea lateral, por lo que tiene una percepción hidrodinámica parecida a los peces. Al salir del agua, después de la metamorfosis, las ranas perciben señales acústicas aéreas, lo que ilustra el uso natural de estructuras anatómicas mediante la adaptación funcional a nuevas necesidades ambientales. Este fenómeno no sólo ocurrió en la evolución natural durante milenios, sino que se repite en la metamorfosis de cada animal, permitiendo su observación directa y sirviendo como un modelo descriptivo o experimental[24].
En los anfibios, aparece, pues, el oído medio. La cóclea presenta una estructura muy simple. El oído inicia su desarrollo a partir de este escalón filogenético, mediante el desarrollo de la lagena que se va alargando, enrollándose progresivamente hasta formar la cóclea o laberinto definitivo en los mamíferos. El oído externo consta solo de una membrana timpánica, en general grande, y situada directamente en la pared exterior a nivel de la piel. El oído medio consiste en una cavidad aérea que es la caja del tímpano en la que un solo huesecillo (columela u osículo) trasmite las vibraciones del tímpano a la ventana oval del oído interno y que desemboca en la faringe por la trompa de Eustaquio.
Todo ello se observa en al Il IV.3.22. En el oído interno, el laberinto se asemeja al de un embrión humano de unos 50 días.
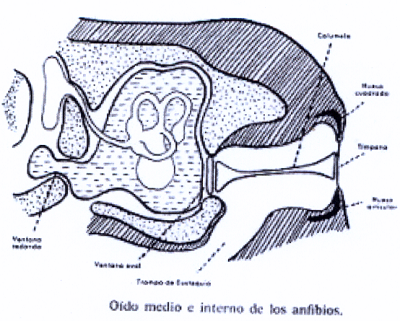
IV.3.22
En algunos urodelos reptantes las vibraciones son apreciadas y transmitidas desde las mandíbulas. En otros, uno de los cuadrúpedos, en los que la mandíbula ha perdido el contacto con el suelo, existe un opérculo que percibe las vibraciones a través de la unión muscular entre la cintura escapular y la ventana oval, por medio de los miembros anteriores.
En los reptiles, el órgano auditivo experimenta aun más cambios evolutivos que en los anfibios, aunque su anatomía en general es parecida, con un pequeño conducto auditivo externo, dos cavidades timpánicas con sendas columelas o columnillas, comunicadas por trompas de Eustaquio que se abren conjuntamente en el techo de la faringe. Pero, en el oído interno, aunque el laberinto membranoso se diferencia poco del de los anfibios, en los reptiles aparece ya el laberinto óseo rodeando al membranoso, primero esponjoso y luego osificado. Al rodear el óseo al membranoso existe un espacio entre los dos con perilinfa y aparece el esbozo de las dos ventanas, oval y redonda, tal como se muestra en la Il IV.3.23.
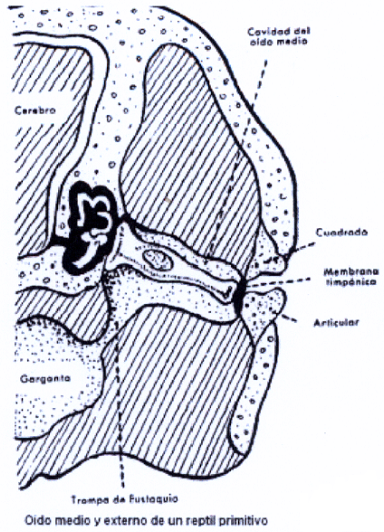
IV.3.23
Otros, sin embargo, no poseen oído medio, como las serpientes; que no tienen cavidad timpánica pero si una columela que avoca al hueso craneal externo y que como los anfibios más primitivos pueden captar las vibraciones del suelo y quizás oír sonidos agudos por su alta energía. Los demás reptiles tienen caja timpánica atravesada por una gran columela.
Estructuralmente, el oído de las aves es similar al de los mamíferos, con oído externo, medio e interno; aunque arrastra algunas peculiaridades propias de los reptiles con aparato auditivo más evolucionado.
Sin embargo, el oído externo carece de pinae, la oreja típica de los mamíferos. En su lugar, las aves tienen plumas auriculares que cubren la apertura del oído y están bajo control muscular, facilitando la penetración del canal auditivo por los sonidos. Las aves acuáticas tienen un repliegue de la piel que se cierra cuando el ave se zambulle, evitando la inundación del oído externo y cualquier daño al oído causado por el aumento rápido de la presión del agua.
En el oído medio, la transmisión de la vibración en la que consiste la onda sonora al oído interno se asegura mediante un solo huesecillo columelar, como en los reptiles que tienen oído medio.
En el oído interno hallamos, también, sáculo y utrículo, con células ciliares que parten de la membrana basilar sumergidas en endolinfa y que, a través del órgano de Corti y el nervio auditivo, alcanzan el encéfalo medio del ave.
Los órganos de los insectos que tienen como objeto captar las ondas sonoras son variados y, por el escaso tamaño del animal, con configuraciones anatómicas bastante originales.
Aunque la inmensa mayoría de los insectos pueden percibir las ondas sonoras, lo hacen mediante cuatro sistemas distintos:
Los ortópteros (saltamontes y grillos), tienen tímpanos, aunque situados en diferentes partes del cuerpo. Situados en el primer segmento torácico en algunas especies de saltamontes y en las patas delanteras de otras y comúnmente en estás en las diferentes especies de grillos.
La anatomía de los oídos alojados en las partes delanteras de los saltamontes Corifora gorgonensis –una especie de la Isla Gorgona del Pacífico que constituye un Parque Nacional Natural de Colombia- fue estudiado por un equipo de la Universidad de Bristol, encontrando un sistema de cierta complejidad y descubriendo una vesícula acústica hasta ahora desconocida y que se explica partiendo de la ilustración Il IV.3.24.
.png)
IV.3.24
Fuente: Investigación y Ciencia, mayo 2014. pág.52
En esta especie, el sonido ambiental que, se capta en los espiráculos torácicos, se amplifica en la traquea acústica –un tubo lleno de aire que deriva del sistema respiratorio y que se origina en el espiráculo torácico- y es conducido a través de ella al tímpano correspondiente; este transfiere las vibraciones a la vesícula auditiva (representada con color amarillo en la Il IV.3.24), por medio de la placa timpánica: una placa de cutícula que realiza la misma función de la columela en el oído medio de los reptiles más evolucionados, acoplando –en este caso- la membrana timpánica con la vesícula auditiva y junto a ella la cresta acústica de –color naranja- donde se encuentran las células receptores, a imagen del órgano de Corti y de los vertebrados). En la cresta se traducen las ondas de presión en sus respectivas frecuencias; y, después, la energía sonora se disipa en el canal de la hemolinfa (de color verde claro)[25].
Los oídos de algunas especies de saltamontes son extraordinariamente sensibles y pueden detectar señales extremas ultrasónicas –más de 130 Khz- a largas distancias.
También tienen tímpanos las cicadas o cigarras (especies Cicadidae y Hemiptera) y algunas familias de Lepidopteros (como Noctuidae, Geomitridae y Pyralidae). Los Noctuidae pueden oír frecuencias de hasta 240 Khz, lo que les permite oír a los murciélagos ecolocalizando sus presas mediante la emisión de chillidos en frecuencias de 20 a 100 Khz. También perciben los insectos los sonidos mediante sensilias auditivas, pelos auditivos que consisten en un mecanosensor con forma de pelo o cerda provisto en su base de una célula nerviosa y una conexión neuronal que también pueden tener otras formas representadas en la Il IV.3.25.
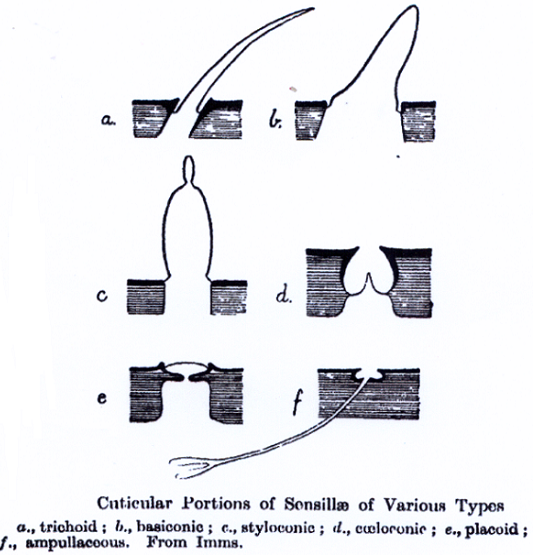
IV.3.25
Las larvas del lepidópteros y ortópteros utilizan pequeños pelos, llamados setas, para sentir las vibraciones sonoras. Las orugas suelen responder a este tipo de sonidos adoptando comportamientos defensivos; tales como permanecer inmóviles o adoptar posturas de lucha.
En los grillos y cucarachas, también este mecanismo de percepción, en forma de pelos “cerci”, recubre parcialmente su abdomen. Son también comunes en las moscas y en todos los insectos peludos.
Un tercer órgano de audición de los insectos presente en las polillas halcón (Cherocampini) es el pilifer labrum en asociación entre este órgano -“el labio que porta pelos”- y el segundo palpo labial, cuya superficie interior esta hinchada por una bolsa de aire concava y de diámetro desigual. El sonido produce vibraciones en este segmento del palpo labial, que se transmiten por contacto a la parte más separada del pilifer que cuenta con un lóbulo pequeño del que parten unos pocos pelos o cerdas y que es un fonoreceptor, como hemos dicho, parte de la base del pilifer unida por una derivación al nervio labral. A través de ese fonoreceptor, los estímulos mecánicos se traducen en estímulos eléctricos que permiten percibir las diferentes frecuencias, receptor acústico que juega el mismo papel que realiza el tímpano en otras polillas o mariposas nocturnas[26].
El órgano de Johnston, presente en las antenas de todos los insectos y que indica directamente la posición y movimiento de estás, indirectamente facilita el mantenimiento de la trayectoria de vuelo en presencia de corrientes de aire variables o turbulentas, en las moscas drosofila y langostas; de la misma forma, durante la natación, a los escarabajos gyrinidae, sirve a los mosquitos machos culicidae como otro órgano de audición –el cuarto, pues, de los insectos- ubicado en el segundo segmento o pedicelo de la entena de ese mosquito, compuesta de tres células sensoras y tres células envolventes. Con ese órgano, el mosquito siente el vuelo de las hembras de su especie para el apareamiento.
El sentido de la audición viene prácticamente siempre enlazado en las especies a la comunicación por sonidos; esto es: a la recepción de señales sonoras producidas por otros miembros de la especie.
Probablemente por esa razón, se explica a continuación Aristóteles sobre la voz de los animales.
Comienza por decir que la voz (φωνὴ) y el sonido (ψόφος) son dos cosas distintas y el lenguaje (δίαλεκτος) es una tercera; que la voz se emite por la laringe, que los animales que no tienen pulmones no tienen voz[27] y que el lenguaje es la articulación de la voz por la lengua: las vocales (φωνήεντα) se emiten por la voz y la laringe, las consonantes (ἄφωνο) por la lengua y los labios. Unas y otras constituyen el lenguaje[28]. Sin embargo, prosigue Aristóteles “es posible emitir sonidos con otros órganos”, “los insectos no respiran” y, por eso, “no tienen ni voz, ni lenguaje”, emiten sonidos “con el aire interior y no con el aire exterior”.
“Los unos zumban, como las abejas y otros insectos alados, plegando y desplegando sus alas, pues el sonido resulta de la fricción con el aire” (πνεύματος). De otros, se dice que cantan, como las cigarras, que emiten sonidos con una membrana que tienen bajo el “hiposoma” –caparazón- cuando lo tienen (διῄρηται[29]). “En cuanto a los grillos (ακρίδες), frotando sus patas traseras (πηδαλίοις) hacen su sonido[30]”.
Respecto a los habitantes del agua, Aristóteles nos dice que ni los cefalópodos ni los crustáceos producen naturalmente ruido (φυσικὸν ψόφον).
En cuanto a los peces, tampoco tienen voz, “pues no tienen ni pulmones, ni traquea, ni laringe”; “pero algunos emiten sonidos, pequeños gritos, que se les llama su voz; por ejemplo la lira[31] y el jromís –estos peces producen una especie de gruñido ahogado- el jabalí del río Aquelous[32], el calcis[33] y el pez cuco[34]. El jromis que Pierre Louis traduce como “l’ombrine”[35] y D’Arcy Thompson como “the sciaena”, deber ser, con toda probabilidad un sciénido (scienidae) cuyas 275 especies, perciformes –esto es: en forma de perca-, designan corvinas, corvas –y, entre ellas, la sciaena umbra u ombrine, que no se encuentra hoy en el Mediterráneo-, verrugatos y roncadores y entre estos últimos, la pagonia cromis o pez tambor (black drum) que produce sonidos entre 100 y 500 Hz, en sus llamadas de apareamiento. Estos sciénidos producen sus sonidos con dos músculos pectorales, largos y estriados, comunes a ambos sexos, que rodean y baten, vibratoriamente la vejiga natatoria. En el Mediterráneo sigue siendo común la corvina argyrosomus regius que incluso se introduce en su estadio juvenil en el agua dulce y emite fuertes sonidos que se oyen a gran distancia.
“Todos estos animales –prosigue Aristóteles- producen lo que parece ser una voz: los unos por frotamiento con las branquias que tienen abundantes aristas; en los otros provienen de las partes internas que rodean el abdomen pues todas encierran aire que se agita y frota para producir los sonidos, y también ciertos selacios[36], parecen lanzar pequeños gritos. Pero no sería exacto decir que estos animales tienen voz, sino solo que emiten sonidos”[37].
Sin embargo:
“El delfín emite, también, un pequeño grito, un murmullo cuando está fuera del agua, al aire libre, aunque no igual (ὁμοίως) a los que acabamos de señalar: pues en el caso del delfín se trata de una voz, pues tiene pulmón y traquea; solo que como su lengua no es libre y no tiene labios, no puede emitir sonidos articulados (ἄρθρον)[38]”
“Entre los animales (reptiles) que tienen lengua y pulmón, los..... ovíparos tienen una voz débil –prosigue Aristóteles-: unos emiten un silbido largo (συριγμόν), como la serpiente; otros tienen una voz débil y quebradiza y otros emiten un débil silbido (σιγμὸν/συριγμόν μικρόν) como las tortugas. La rana tiene una lengua de forma excepcional: la parte frontal, que en otros animales (cuadrupedos terrestres) pueden moverse libremente, está fijada (al paladar) como en los peces; pero la parte junto a la laringe esta libre y replegada, lo que permite a la rana lanzar el grito que la caracteriza
El croar de las ranas en el agua es el grito de los machos que invitan a la hembra al acoplamiento (ὸχείαν); pues cada animal tiene un grito particular a ese efecto.
El macho de la rana produce el suyo poniendo al ras del agua su mandíbula inferior y distendiendo la superior. Sus sacos bucales en sus mejillas se vuelven transparentes bajo el efecto de la tensión y parece que sus ojos brillan como lámparas a través de la piel; pues la copula se produce generalmente por la noche[39]
La clase (γένος) de las aves también emiten gritos (φωυήν); los que tienen la capacidad de pronunciar palabras (δίάλεκτον) son los que tienen la lengua larga, plana y fina. En algunas especies, hembras y machos lanzan los mismos gritos; en otras, son distintos. Los pájaros pequeños tienen cantos más variados y trinan más que los grandes, pero es en la época del apareamiento (ὀχείαν) cuando cada especie se pone a cantar. Hay otros pájaros que gritan cuando pelean -como la codorniz (ὄρτυξ)-; otros cuando desafían al adversario –como las perdices- (πέρδικες) –o cuando vencen como los gallos de corral (ἀλεκτρυόνες)-. En algunas especies tanto los machos como las hembras cantan, como es el caso de los ruiseñores; sin embargo, la hembra deja de cantar cuando incuba su puesta; en otras especies, por el contrario, el macho canta y las hembras no, como es el caso de las codornices y las gallinas[40].
Entre las especies de aves, dependiendo de su diversidad de origen, las mismas especies pueden tener cantos distintos; por ejemplo: algunas perdices cloquean (κακκαβίζουσιν) otras pían (τρίζουσιν)[41]”.
Igualmente, mantiene Aristóteles que puede diferir el canto de los padres y las crías si no han crecido entre ellos, sino entre otros pájaros y que incluso se ha visto a un ruiseñor enseñar a cantar a un pájaro más pequeño de una especie distinta, lo que prueba que el canto puede formarse por la educación.[42]
“Los mamíferos tienen distintas voces, pero ninguno de ellos pronuncia palabras lo que está reservado al hombre, y no a todos los hombres: los sordos de nacimiento son también mudos; emiten sonidos, pero no pronuncian palabras. Los niños, mientras no tienen el control de las demás partes de su cuerpo, tampoco controlan su lengua, que no se libera eficientemente más que más tarde; hasta entonces cecean y tartamudean.”
Efectivamente, como dice Aristóteles, los peces emiten sonidos y el hidrofono que permite escuchar dentro del agua esos sonidos ha venido a confirmar y ampliar las observaciones del estagirita tanto en relación a la producción efectiva de sonido como a la forma de producirlo, a su frecuente asociación con la reproducción sexual; de hecho, el conjunto de observaciones en la naturaleza que efectúa Aristóteles es tan importante que, incluso ahora, hay algunas que no se han comprobado
Sin embargo, se equivoca afirmando que los insectos no respiran. Como todos los seres vivos, los insectos respiran inhalando oxigeno del aire y exhalando dióxido de carbono; aunque a diferencia de la mayoría de los vertebrados de respiración aérea, los insectos terrestres a los que se refiere Aristóteles, como invertebrados, carecen de pulmones y de sangre (la hemolinfa realiza otras funciones) y obtienen el oxígeno del aire por medio de una red de conductos (traqueas) que se abren al exterior a través de orificios respiratorios (estigmas) y que pueden cerrar o ocluir mediante espiráculos u ostiolos. Las traqueas se ramifican en traqueolas microscopicas, cuyas paredes son finas y permeables. Y por difusión a través de esas paredes, según la concentración relativa de los gases a ambos lados de la membrana que tiende a igualarse, llega el oxígeno a todos los órganos del cuerpo del insecto y se evacua el dióxido de carbono.
Pero es cierto que los insectos no producen sus cantos o sonidos impulsando el aire de su respiración. El modo natural de producir esos cantos o sonidos es mediante el frotamiento de ciertas partes del cuerpo o estridulacion, técnica que utilizan también algunas aves o reptiles. Estos cantos tienen como objeto, muy frecuentemente, la comunicación. Los zumbidos a que también se refiere Aristóteles los emiten accidentalmente los insectos de vuelo rápido como las moscas y moscardones.
El estruendoso canto de la cigarra lo producen los machos de la especie con las membranas vibratorias quitinosas de su abdomen, llamadas timbales y de sacos aéreos que sirven de caja de resonancia, ubicados todos ellos en el costado del primer segmento abdominal. Con tal intensidad que pueden llegar a morir por la presión producida por sus ondas acústicas que alcanzan hasta 100 decibelios a un metro de distancia. Tiene ese canto el propósito de atraer a las hembras y mostrar su excitación en caso de peligro.
El canto de los grillos, muy variado en sus características y periodo del año en que se emite, corresponde a parecidas circunstancias a las de las cigarras.
Sin embargo, el grillo no lo produce con sus patas traseras, como dice Aristóteles. El órgano de estridulacion son los elitros o alas delanteras quitimizadas, cruzadas por una arista ancha que discurre por el centro de cada elitro con un filo o borde en forma de sierra; mientras que el borde trasero del elitro termina ásperamente. El grillo, rítmicamente, alza los dos elitros, raspando con el borde de uno la arista en forma de sierra del otro, y el sonido se amplifica resonando en el elitro cruzado por la arista en forma de sierra, en el caparazón superior y hasta en su madriguera, como vemos en la ilustración Il IV.3.26[43].

IV.3.26
Dentro de la variedad inherente a los cantos de las diferentes especies de grillos, también en cada especie varia el repertorio. El canto de llamada, bastante alto, atrae a las hembras y repele a otros machos. Usa el canto de cortejo, más discreto, cuando la hembra se aproxima, y el canto triunfal, por un periodo breve después de la cópula. Por el contrario, un canto más fuerte y acelerado se dirige a otro grillo macho cuando detecta su presencia.
La temperatura ambiente acelera también el ritmo del canto; hasta el punto de que según la Ley de Dolbear, el número de chirridos producidos en 14 segundos por el grillo arboreo de las nieves –común en Estados Unidos- más 40, arroja aproximadamente la temperatura en grados Farenhait.
Los peces, anfibios y mamíferos que viven en el agua, producen ruidos como señala Aristóteles.
Entre ellos se encuentran, efectivamente, la familia de los trigleidos que hacen vibrar su vejiga natatoria.
También es acertado, lo que señala Aristóteles respecto al delfín: como tiene pulmones, traquea y boca, puede lanzar un grito, que es una voz. Lo mismo por cierto que los mamíferos que viven en el mar, que emiten un grito armónico, con una gran potencia y alcance dentro del medio acuático, tal como las ballenas y las orcas.
Esos mamíferos marinos, probablemente son capaces de introducir en su voz combinaciones significativas; porque son capaces de comprenderlas.
El grupo de Louis Herman del acuario Kewalo en Honolulu, diseñó en los años ochenta del siglo pasado dos lenguajes artificiales que enseñó a los delfines mulares del acuario, lenguajes de señas o gestos correspondientes a palabras que podían componerse en frases; de forma que los delfines entendían y ejecutaban las frases con sentido, mientras permanecían pasivos cuando no lo tenían[44].
En términos generales, la única percepción de Aristóteles no confirmada es la capacidad de los seláceos (σελαχῶν) de emitir sonidos.
Como hemos dicho, mediante el hidrofono, y mediante las observaciones en acuario, se han detectado otras especies de animales acuáticos que producen sonidos: a) los hipocampos o caballitos de mar (genero Hippocampus) que emiten un pequeño clic o chasquido producido por el choque o roce de su cráneo contra las placas óseas de su cuello; b) los balistes o peces luna que emiten chillidos y crujidos con su dientecillos de la faringe; c) las chicharras o golondrinas de mar (cephalacanthus volitans) que emiten quejidos estridentes al frotar las espinas de su cuerpo quitimizado, ampliando el sonido con su vejiga natatoria[45]
Fuera de la zona mediterránea, los peces sapo (Batrachoididae), peces bentonicos, que también, emiten zumbidos en la época de apareamiento, de baja frecuencia, pero de intensidad tal que se perciben dentro de las casas o a la orilla del mar, y sonidos guturales que corresponden a circuitos neuronales similares a los vertebrados[46] con ciertos patrones verbales –zumbidos para llamar a la hembra, gruñidos para enfrentarse a los adversarios- y escuchan durante milisegundos en los periodos en que no producen sonido.
Otro animal, fuera de la cuenca mediterránea, que produce un sonido significativo, es el camarón pistola o camarón arado (de la familia Alpheidae) que se defiende y aturde a sus presas mediante el empleo de una pinza modificada que produce un fuerte chasquido sónico[47]
Entre los animales de sangre fría -poitilotermos- solo algunos teleósteos o peces de esqueleto óseo, reptiles o anuros son capaces de producir sonidos.
Los teleósteos producen sonidos, en una secuencia de golpes que ocasionan golpeando con el cuerpo o mediante la expulsión de aire de la vejiga natatoria, también como medio de comunicación sexual. Los reptiles mediante órganos específicos, tales como colas corneas, resonantes y expulsión sibilante de aire.
Los anuros –los sapos y las ranas- son, como todo el mundo sabe, los tenores anfibios de las noches tranquilas, utilizando para su poderoso croar sus sacos bucales de resonancia, externos los de los sapos.
Los vertebrados terrestres, producen los sonidos mediante laringes y cuerdas vocales, impulsados por el aire que utilizan para la respiración. Las aves y los cocodrilos, mediante la siringe, un conducto tubular cartilaginoso ubicado en la bifurcación de la traquea que conduce a los bronquios el aire de la respiración; hasta el punto que los pájaros que no tienen siringe –como el buitre americano- son mudos.
Al menos en los vertebrados, el sonido significativo viene asociado al lenguaje gestual. En el camino de montaña, la vaca se vuelve hacia su becerro que vacila o se distrae y emite un mugido corto y fuerte para animarle; el león ruge y adopta una actitud amenazadora mostrando sus colmillos, cuando desea atemorizar a un oponente; el halcón grita reiteradamente y se cierne sobre la cabeza del intruso que se acerca a su nido; en todos estos casos el sonido unido a la acción son suficientemente elocuentes.
Pero, además, otros animales son capaces de introducir en su voz más contenidos o de comprender combinaciones significativas. Ya hemos hablado de los delfines.
La primatologa Sue Savage-Rumbaugh adiestro a Kanzi, un macho de bonobo, a comunicarse tocando símbolos de un tablero de símbolos léxicos y a comprender el lenguaje hablado. La primatologa estimaba que Kanzi podía seleccionar 300 palabras coherentes y comprender más de 1000 palabras habladas.
El trabajo de Savage-Rumbaugh y de otros investigadores, sugiere razonablemente que los simios podían comprender y emplear los conceptos a los que hacen referencia los símbolos lógicos y utilizar palabras mediante símbolos para representar cosas de su alrededor; así como combinar esas palabras para hacer peticiones, dar información y realizar observaciones sobre su entorno. Bearzi y Stanford entendían que el nivel de entendimiento de Kanzi era similar al de un niño de un año[48]
La Il IV.3.27 muestra a Kanzi “leyendo” junto a su tabla de símbolos léxicos.

IV 3.27
Caso evidente de la voz significativa es la voz humana.
La voz es, evidentemente, el sonido producido por un instrumento formado por una fuente de energía, (los pulmones) un oscilador, (los ligamentos o cuerdas vocales) y un resonador (la cavidad vocal).
La principal función de los pulmones es producir una sobrepresión de aire generando así un chorro de aire en los otros dos elementos del instrumento. El aire pasa por la glotis, que es un espacio abierto en la base de la laringe entre los dos ligamentos verticales vocales (denominados, usualmente, cuerdas vocales, auque se trata de repliegues elásticos de la mucosa que recubre la laringe). El extremo delantero de cada ligamento vocal se halla sujeto al cartílago tiroides o nuez de Adán –así llamado por ser característicamente más prominente en el género masculino-. El extremo opuesto, por tanto trasero, se halla sujeto a uno de los pequeños cartílagos aritenoides; estos son móviles, y se desplazan hasta conseguir la separación de los ligamentos (en la respiración), o su acercamiento y extensión. Además de generar sonidos, los ligamentos vocales tienen otra misión; proteger a los pulmones de la penetración de pequeños corpúsculos arrastrados en el flujo del aire inspirado. Situados sobre estos ligamentos o cuerdas vocales se encuentran los dos ligamentos vocales “falsos” que operan cerrando la respiración con una sobrepresión de aire en los pulmones. Estos ligamentos, se encuentran en el fondo de la laringe, cavidad más amplia, de forma tubular, acoplada a la faringe, que une la boca con el esófago. El techo de la faringe está formado por el velo del paladar o paladar blando que, a su vez, franquea la entrada a la cavidad nasal, cuando el velo del paladar está en posición alzada, es decir, cuando se pronuncian las vocales –pero no las nasales-, se cierra el paso a la nariz y el aire sale por la boca. Laringe, faringe y boca, conjuntamente, constituyen el conducto o tracto vocal, que es una cámara de resonancia cuya geometría se modifica mediante la posición de la lengua, el paladar, la úvula o campanilla y los labios.
En la Il IV.3.28 se muestra la disposición de estos órganos.
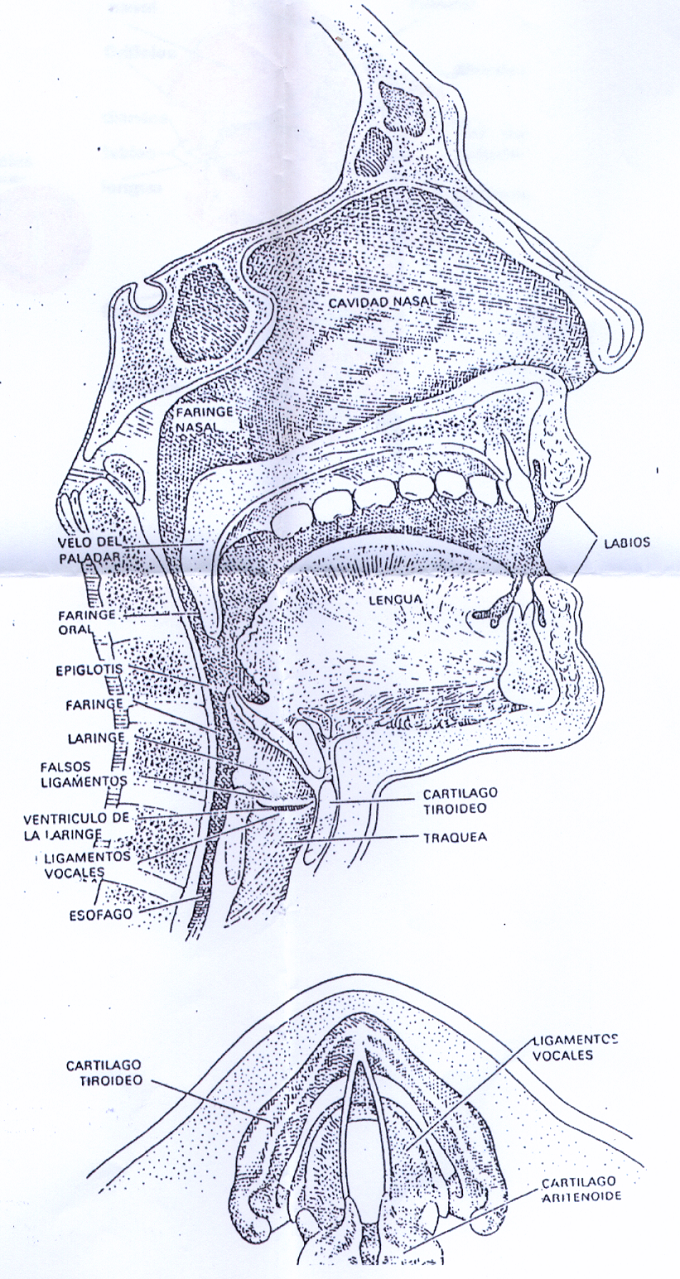
IV.3.28
Hemos visto que son sonidos simples aquellos que tienen una sola frecuencia y, por consecuencia, un solo armónico: caso típico del silbido. No existe, en este caso, ni resonancia, ni actividad de las cuerdas vocales; se produce simplemente la expulsión del aire por los labios, por lo tanto, el sonido se modifica de acuerdo con la apertura del propio órgano; no tiene más complicaciones: tiene un solo armónico y aparece como una sinusoide en la representación gráfica de la onda sonora
Frente a estos sonidos simples nos encontramos con los sonidos complejos, cuando el sonido producido por la fuente sufre resonancias.
En el caso de la voz humana, el elemento de fonación simple son las cuerdas vocales y la caja de resonancia es la cavidad vocal, formada por el conjunto de estructuras fijas y móviles entre la glotis y los labios
El sonido generado por el flujo de aire, entrecortado por la vibración de los ligamentos vocales, se denomina fuente vocal. Se trata, como he dicho, del instrumento del habla o del canto que produce un sonido complejo, formado por una frecuencia fundamental (fijada por la frecuencia de vibración de los ligamentos vocales) y de un gran número de armónicos o sobretonos. La amplitud de los armónicos decrece uniformemente con la frecuencia, en una proporción de unos doce decibelios (12 Db) por octava.
El tracto vocal es, pues, un resonador acústico. La transmisión de un sonido a través de un resonador acústico es función de la frecuencia. Los sonidos correspondientes a la frecuencia de resonancia propia de cada resonador son menos atenuados que otros sonidos y, por lo tanto, son emitidos con una mayor amplitud relativa o con un mayor volumen relativo que otros sonidos; de forma que, cuanto mayor sea la diferencia de frecuencia entre un sonido y su resonancia, más débilmente se percibirá esta. El conducto vocal tiene cuatro o cinco resonancias importantes, denominadas formantes. Por tal razón, todos los armónicos de la fuente vocal pasan por el conducto vocal con más o menos éxito, según su frecuencia; cuanto más cerca esté un armónico de la frecuencia de un formante, tanto más aumentara la fuerza de su señal en los labios.
Las frecuencias de los formantes dependen de la forma del conducto. Suponiendo que el conducto vocal fuera un cilindro perfecto, cerrado en la glotis y abierto en los labios, de 17,5 cm de longitud, lo que resulta bastante verosímil en el caso de un varón adulto, los primeros cuatro formantes tendrían frecuencias cercanas a 500, 1.500, 2.500 y 3.500 Hz, -recordemos que son ciclos por segundo-; si el conducto vocal fuera más corto, estas frecuencias base resultarían distintas; normalmente algo inferiores
El formante más grave corresponde a un cuarto de longitud de onda, lo que equivale a decir que un cuarto (1/4) de la longitud de la onda sonora emitida por la fuente ha cabido en el conducto vocal. Del mismo modo, el segundo, el tercer y el cuarto formante, corresponden, respectivamente, a tres cuartos (3/4) de la longitud de la onda sonora emitida, a uno y un cuarto (1 1/4) y a uno y tres cuartos (1 3/4) de esa longitud de onda.
La apertura de la mandíbula amplia el conducto vocal en la región de los labios y lo reduce en la zona cercana a la glotis; tal apertura resulta decisiva para la frecuencia del primer formante, que aumenta a medida que se va abriendo la mandíbula. La frecuencia del segundo formante es muy sensible a la forma del cuerpo de la lengua; la frecuencia del tercer formante es sensible a la posición de la punta de la lengua. Si movemos estos articuladores de manera diferente, se modifica la frecuencia de los dos formantes más graves (el primero y el segundo), a lo largo de una escala bastante amplia. A cada configuración de articuladores corresponde a un juego de formantes que, a su vez, están directamente asociados con el sonido de una vocal.
En resumen, la modificación de la geometría del tracto vocal significa, favorecer el nivel de la señales fónicas cuya longitud de onda coincide con las fracciones enteras de la cámara de resonancia así conformada que he señalado. Las figuras Il IV.3.29 y Il IV.3.30 muestran la representación grafica de las ondas correspondientes a la emisión vocal de las cinco vocales castellanas por un varón y una mujer normal, en la que se observa el rasgo glótico y el rasgo del tracto vocal y, por tanto, la relación entre la geometría de ese tracto vocal y los diferentes formantes. La señal se ha convertido, mediante el micrófono, en una señal eléctrica que, antes de su transformación o análisis se denomina señal temporal y se registra mediante un osciloscopio o aparato similar, como un espectrógrafo. En las dos ilustraciones anteriores, se representa la intensidad de dicha señal en milivoltios y la de la duración y la de la duración en milisegundos. De esa representación gráfica de la señal temporal se deduce la frecuencia, esto es: el número de periodos o ciclos de la misma por unidad de tiempo. El periodo se computa por los máximos de la señal temporal –esto es: de los “picos” que aparecen en el gráfico- auque no puede hablarse con propiedad de “picos”, -más que en caso de análisis-; la distancia es mayor entre ellos cuanto más grave es la voz; por lo tanto, cuanto más grave es la voz menor es su frecuencia y viceversa, cuanto más aguda, menor es la distancia entre los máximos de la señal temporal y mayor es su frecuencia. Por eso, la mayor frecuencia que observamos en la señal temporal de la voz femenina obedece a ser más aguda que la masculina.
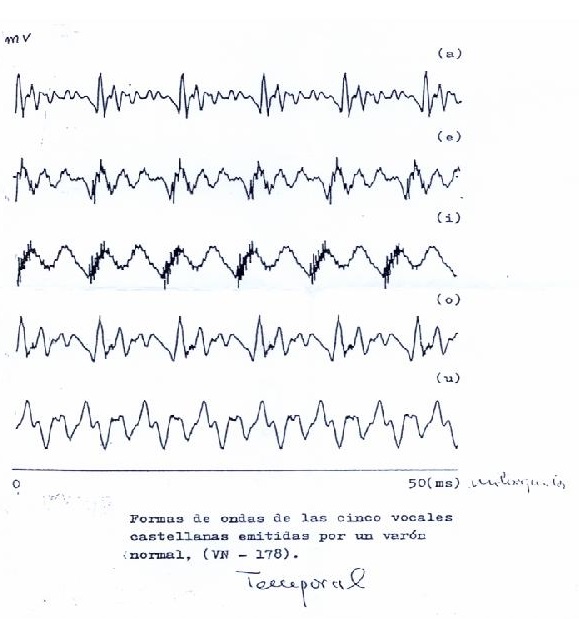
IV.3.29
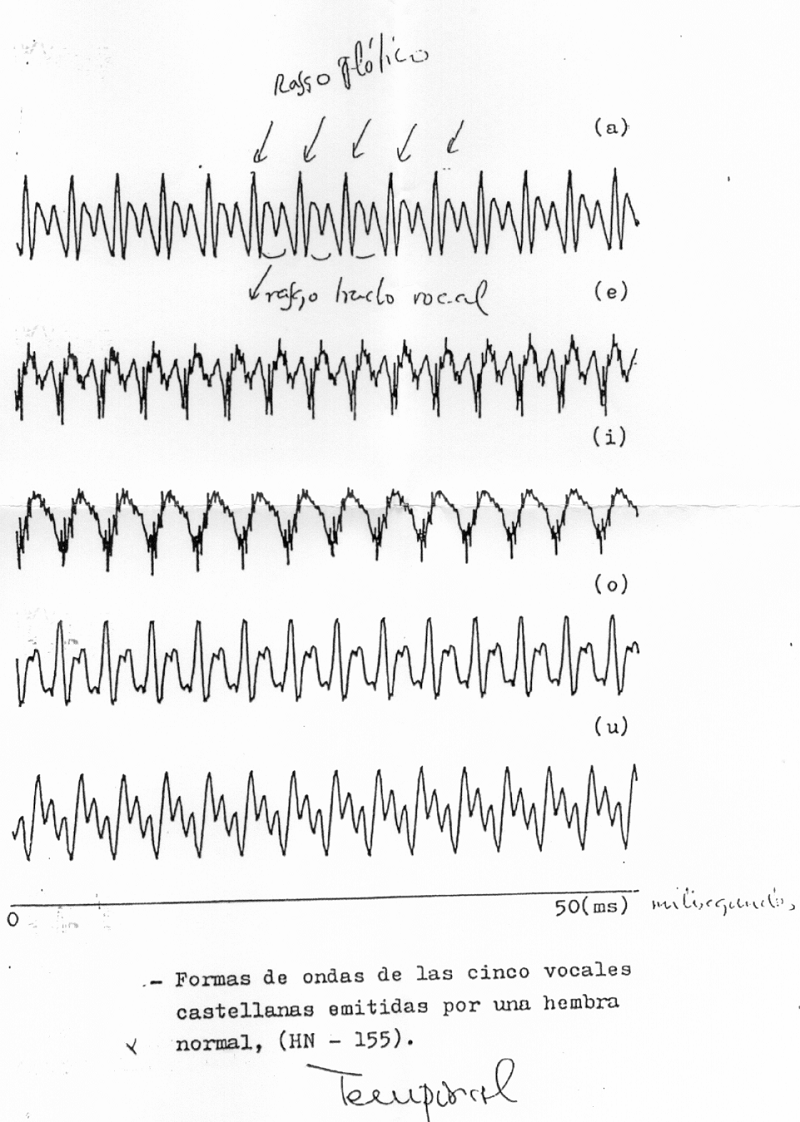
IV.3.30
En la representación grafica de la voz del hombre y la mujer se producen los dos tipos de señales: una, correspondiente a las vibraciones producidas por la glotis –rasgo glótico- y otra, a las resonancias de esas vibraciones en el tracto vocal –los formantes-; que es el rasgo del tracto vocal.
En el espectro producido por la fonación aparecen tanto los armónicos o los grupos de armónicos de la frecuencia fundamental –esto es, el sonido producido por la glotis- como las resonancias de la caja, cavidad o tracto vocal, que dependen de los rasgos anatómicos del hablante; por lo que es evidente, que el modo más eficaz de identificar una determinada fonación con su autor consiste en analizar estas resonancias producidas por el tracto vocal que corresponden a la geometría anatómica de la caja vocal; esto es, a sus rasgos anatómicos diferenciados; detectando en ese análisis las estructuras periódicas en el espectro mediante la transformación inversa[49] y la transformación de Fourier, discriminando la frecuencia fundamental y analizando la resonancia del tracto vocal, los llamados formantes que denotan la particular anatomía del tracto vocal del hablante en la frecuencia típica de 50 y 5.000 Hz.
A la vista de todo lo anterior ya podemos ponderar cuanto de cierto y qué poco de equivocadas tienen las apreciaciones de Aristóteles sobre el sentido auditivo y la voz de los animales.
[1] LC 33, 419b, 10
[2] LC 33, 419b, 17
[3] De hecho el agua es mejor transmisor
[4] LC 33, 419b, 18. Lo que parece que intuye el carácter ondular del sonido, como cuando la piedra cae en el agua y produce ondas. Impresión que se refuerza cuando, más tarde, equipara los efectos del sonido y los de la “repercusión” o reflexión (ἀνάκλασις).
[5] LC 33, 419b, 35.
[6] LC 33, 420a, 4
[7] LC 33, 420b, 19
[8] LC 33, 420b, 30
[9] LC 33, 421a, 1
[10] LC 35, 533a, 20
[11] LC 35, 533a, 30
[12] Ibidem, 533b, 7
[13] Ibidem, 533b, 9
[14] Ibidem, 533b, 33
[15] Labrax Lupus
[16] LC 33, 436b, 10 y LC 32, 980a 21
[17] Lo que es común a la propagación de todas las ondas, incluso las lumínicas, que pueden relantizarse extraordinariamente en función del medio de propagación.
[18] En el que la magnitud básica es el mol, la unidad con la que se mide la cantidad de moléculas de una sustancia, equivalente a 12 gramos no enlazados molecularmente de carbono-12 y a 6,02214129 x 1023 moléculas. Experimentalmente, se ha comprobado que el volumen que ocupa un mol de cualquier gas, es 22,4 L a 0º y a 1.015 mb; a este volumen se le llama volumen molar del gas. El volumen molar es, pues, un cubo cuyos lados miden, más a menos, 28,2 cm.
[19] Puesto que no coincidirá con el valor gamma de otros gases mezclados, ni con los isótopos distintos del gas en cuestión.
[20] Esto es: la recta perpendicular al frente de onda.
[21] Que provienen de la obra, ya abundantemente citada, Life, The Science of Biology, Sunderland (Mass. 1997, pag. 938, como asimismo la descripción funcional de esos diagramas).
[22] Peces muy primitivos que viven exclusivamente en los lagos y ríos de África.
[23] La audición de los peces www.oya-es.net/reportajes/peces/htm
[24] RC 32, Págs. 86-88
[25] RC 33, pág. 52
[26] LC 38, pág. 297.
[27] LC 18, IV. 9 535a
[28] Ob. Cit. IV.9 535a
[29] Esto es, cuando el cuerpo esta compuesto de un soma y un hiposoma –caparazón-
[30] Ibidem, 535b
[31] Se trata del garneo, rubio o arete (trigla lyra); un trigleido, pez bentónico –es decir que vive en el fondo marino- alimentándose de las piezas que desentierra de él y que produce, efectivamente, el gruñido al que se refiere Aristóteles; hasta el punto de que prácticamente no se calla, pues son muy “parlanchines””.
[32] Según D’Arcy Thompson, puede ser un siluro (silurus glanis); esto es: un anfibio, perteneciente a una familia que se distingue por su capacidad vocal.
[33] D’Arcy Thompson, en nota a H.A. 536b 18, recuerda que este pez se ha identificado por mucho tiempo con el pez de San Pedro (Zeus faber) un pez pelagico de aguas profundas.
[34] Otro trigleido, identificado por Linneo como Trygla cuculus; y, en cualquier caso, denominado con el mismo nombre también, hoy, en el Mediterráneo, como observa D’Arcy Thompson en nota a H.A 535, b19.
[35] En su traducción de H.A. 535, b.18
[36] O peces cartilaginosos, por oposición a los teleósteos.
[37] LC 18, 535b, 13-23
[38] Ob. Cit. 536a
[39] LC 18, 536 a, 4-20
[40] LC 18, 536.a, 21-32
[41] En el mismo sentido Teofrasto, fragmento 181 de Teubner; Aelian III, 35; Antigono, De Mirabilia VI; D’Arcy Thompson señala, además, que se trata de una diferencia entre dos especies distintas: Perdrix graeca y Perdrix cinerea, en nota H.A IV.9.536 b, 13 y 14.
[42] LC 018, 536 b, 11-15. D’Arcy Thompson, en nota al mismo pasaje, cita a Warde Fowler (Summer Studies) que mantiene opinión contraria a la de Aristóteles, aduciendo el caso del cuco, ya que la voz innata del cuco no puede modularse como el canto modulado y diestro del ruiseñor.
[43] Wikipedia sv “cricket”
[44] RC 7, pág. 74.
[46] Andrew Baes. Science x network/fish.org/news/2005-07-fish.html
[47] http://1024.drpez.com/julio_05/img_amantis_13.jpg.
[48] RC.7, pág. 74.
[49] Bogert, Healy y Tukey, 1963
{kind=link}